

41) Odbiór i przekazywanie sygnału u roślin - jest to proces odbierania sygnałów docierających do komórek roślinnych ze środowiska zewnętrznego lub wnętrz organizmu. Bodźcem środowiskowym mogą być czynniki abiotyczne, takie jak światło, temperatura, grawitacja, dotyk, substancje chemiczne (woda, CO2, sole mineralne) oraz czynniki biotyczne takie jak patogeny czy sąsiedztwo innych roślin. Czynnikami wywołującymi reakcje komórki są fitohormony, endogenne regulatory wzrostu i rozwoju roślin, siły mechaniczne (turgor) oraz substancje odżywcze albo jony.
Odbiór bodźca: Odbiór bodźca jest możliwy dzięki obecności struktur określanych jako receptory. Receptory roślinne dzielone są na receptory błonowe i wewnątrzkomórkowe. Receptory błonowe są związane z błoną komórkową. Mogą znajdować się na jej powierzchni lub wewnątrz błony. Receptory wewnątrzkomórkowe mogą znajdować się w dowolnym kompartmencie komórki. Lokalizacja receptorów decyduje o zakresie odbieranych bodźców. Receptory błonowe mogą odbierać informacje, które nie wpływają bezpośrednio na działanie komórki. Zapewniają one między innymi reakcję na pojawiające się w otoczeniu komórki substancje chemiczne, które nie mają możliwości przejścia przez błonę półprzepuszczalną. Receptory wewnątrzkomórkowe są odpowiedzialne za odbiór bodźców wnikających do komórki. Bodźcem odbieranym przez receptory wewnątrzkomórkowe mogą być czynniki fizyczne (np. światło), a także cząsteczki chemiczne wnikające do wnętrza komórki. Dotychczas dobrze poznano budowę molekularną kilku roślinnych fotoreceptorów. Udowodnione zostało także istnienie szeregu receptorów uczestniczących w reakcjach hormonalnych. Analiza zsekwencjonowanego genomu Arabidopsis thaliana pozwoliła ustalić, że zawiera on geny ponad 340 kinaz mogących spełniać funkcję receptora (RLK - ang. receptor like kinase).
TRANSDUKCJA SYGNAŁU: Transdukcja, łańcuch przekazywania sygnału, jest procesem przekazania informacji z receptora do miejsca docelowego. W efekcie możliwa jest rakcja komórki na docierający bodziec. W komórkach roślinnych występują trzy typy układów transdukcyjnych: jednoskładnikowe, dwuskładnikowe i trójskładnikowe. Układ jednoskładnikowy składa się z pojedynczej cząsteczki lub kompleksu integralnych białek błonowych. Domena wystająca poza błonę komórkową pełni funkcję receptora łączącego się z ligandami. Domena znajdująca się po stronie cytoplazmatycznej ma właściwości specyficznej kinazy tyrozynowej. Receptory tego typu zostały dobrze poznane w komórkach zwierzęcych i są nazywane receptorową kinazą tyrozynową (RTK). W komórkach roślinnych wykryto jedynie geny kodujące białka o podobnej strukturze. Układy transdukcyjne dwuskładnikowe składają się z dwóch białek. Pierwsze z nich, nazywane jest sensorem, składa się z dwóch domen. Zwykle domena N-końcowa pełni funkcję receptorową, a domena C - końcowa jest kinazą histydynową. Receptory o tego typu odbierają sygnał w postaci gazowego fitohormonu - etylenu. W przypadku receptorów błonowych domena będąca sensorem wystaje z błony komórkowej. Białka tego typu mogą być również białkami rozpuszczalnymi. Przyłączenie liganda receptorowej prowadzi do autofosforylacji domeny C - końcowej. Grupa fosforanowa jest następnie przenoszona na drugi element układu transdukcyjnego nazywany białkiem regulatorowym. Reszta fosforanowa pozostaje połączona z resztą histydynową, aby następnie zostać przekazana na asparaginę regulatora odpowiedzi. To białko oddziałuje bezpośrednio z odcinkami promotorowymi właściwych genów.
Układy transdukcyjne trójskładnikowe zawierają receptor oddziałujący z białkiem sprzęgającym. Oba elementy znajdują się w plazmolemmie. Białko sprzęgające aktywuje enzymy lub kanały jonowe określane jako efektory. Efektem jest zmiana stężenia wtórnego przekaźnika informacji. Związki pełniące funkcję wtórnego przekaźnika wpływają zaś na aktywność enzymów lub na inne białka o charakterze efektorowym. W układzie trójskładnikowym indukcja receptora jest powiązana ze wzrostem aktywności kinazy serynowo-treoninowej.
WTÓRNE PRZEKAŹNIKI INFORMACJI: Występujące w układach trójskładnikowych wtórne przekaźniki informacji w komórkach eukariotycznych pojawiają się w wyniku działania białka G. U Arabidopsis thaliana został wykryty gen GPA1, który koduje podjednostkę alfa białka G. Polipeptyd zbudowany jest z 383 reszt aminokwasowych i ma masę 44,6 kDa. Chociaż informacje o obecności i budowie roślinnego białka G są niepełne, wydaje się, że może ono funkcjonować w formie trimerycznej tak jak ma to miejsce u zwierząt. Heterotrimeryczne formy białka G uczestniczą w przekazywaniu sygnału pochodzącego od auksyn, giberelin, cytokinin oraz fitochromu i kryptochromu. Poza białkami G tego rodzaju, u roślin stwierdzono występowanie małych białek G z rodziny RHO.
Monomeryczne białka G uczestniczą w przekazywaniu sygnału pochodzącego od kwasu abscysynowego i brassinosteroidów. Białko G oddziałując z kompleksem ligand-receptor przyłącza resztę fosforanową do GDP związanego z podjednostką alfa. Następnie aktywowana jest fosfolipaza C (PLC), która katalizuje reakcję rozpadu fosfatydyloinozytolobisfosforanu (PIP2). Produktami reakcji są dwa związki uznawane za wtórne przekaźniki informacji: inozytolo-(1,4,5)-trifosforan (IP3) i 1,2-diacyloglicerol (DAG). IP3 jest związkiem rozpuszczalnym, przemieszczającym się w cytoplazmie. Zwiększone stężenie IP3 powoduje otwarcie kanałów wapniowych obecnych w strukturach cytoplazmatycznych i wypływ jonów Ca2+ do cytozolu. DAG jest związkiem hydrofobowym, pozostaje więc w obrębie błony komórkowej powodując aktywację kinazy białkowej C. Podwyższone stężenie jonów Ca2+ w cytozolu jest bezpośrednią przyczyną aktywacji wielu enzymów. Przy braku pobudzenia stężenia jonów wapniowych w cytozolu jest bardzo niskie (około 100-200 nmol/l-1). Niemal cała pula Ca2+ zgromadzona jest w magazynach takich jak cysterny siateczki śródplazmatycznej i sarkoplazmatycznej, wakuoli i innych organellach oraz apoplaście. Pojawienie się IP3 prowadzi do wzrostu stężenia jonów wapnia do około 1 1 μmol l-1. Kanały wapniowe mogą być wrażliwe na IP3 lub otwierać się pod wpływem innego czynnika. Znane są trzy rodzaje kanałów wapniowych: kanały reagujące na zmiany potencjału elektrycznego plazmolemy (typ VOC - ang. voltage operated channels), kanały reagujące na przyłączenie cząsteczki sygnałowej (typ ROC - ang. receptor operated channels) i kanały reagujące na zmiany turgoru (typ SOC - ang. streach operated channels). Receptory odbierające bodźce środowiskowe mogą współdziałać nie tylko z białkiem G lecz także wywoływać zmiany potencjału błonowego. Reakcja o takim charakterze zachodzi w wyniku odebrania bodźca mechanicznego i mimozy (Mimosa pudica). Po pobudzeniu i gwałtownym wypływie jonów wapnia do cytozolu dochodzi do wolniejszego ich powrotu do magazynów w yniku działania pomp wapniowych. Do aktywacji enzymów związanej ze wzrostem stężenia Ca2+ dochodzi przy udziale kilku białek łączących się z jonami. Najlepiej poznanym białkiem wiążącym jony wapnia jest kalmodulina. Jej obecność została potwierdzona u wszystkich przebadanych gatunków roślin. Aktywna kalmodulina łączy się z białkami docelowymi, którymi mogą być enzymy, pompy wapniowe, kinazy i fosfatazy białkowe oraz białka cytoszkieletu. Podstawowym mechanizmem zapewniającym odpowiedź na docierający do receptorów bodziec jest fosforylacja i defosforylacja białek. Kinazy białek przenoszą resztę fosforanową z ATP na białko docelowe, zaś fosfatazy katalizują hydrolityczne oderwanie reszty fosforanowej. Genom Arabidopsis thaliana zawiera ponad 100 genów kodujących kinazy białkowe. Poza kinazami aktywowanymi przez jony wapnia i fosfolipidy stwierdzono obecność kinaz zależnych od cyklicznych nukleotydów takich jak cAMP i cGMP. Powstanie i rozpad tych związków jest możliwy dzięki enzymom określanym jako cyklazy i fosfodwuestrazy. Chociaż obecność obu cyklicznych nukleotydów została stwierdzona w komórkach roślin oraz znane są procesy przez nie regulowane brak jest jednoznacznych dowodów na istnienie enzymów odpowiedzialnych za syntezę tych związków. Zostąło jednak dowiedzione, że cGMP uczestniczy w regulacji syntezy amylaz w warstwie aleuronowej ziarniaków zbóż indukowanej przez gibereliny oraz w regulacji wytwarzania białek wiążących chlorofil i enzymu Rubisco w chloroplastach. Drugi z cyklicznych nukleotydów, cAMP uczestniczy w regulacji syntezy antocyjanów.
Za wtórne przekaźnik informacji uznawany jest także tlenek azotu (NO). Związek ten uczestniczy w komórkach roślinnych w odpowiedzi na atak patogenów, procesie dojrzewania i starzenia się. Funkcję wtórnego przekaźnika sygnału mogą także spełniać produkty rozpadu sfingolipidów. Fosforan sfingozyny jest wytwarzane podczas reakcji na kwas abscysynowy.
42) Odporność roślin na czynniki środowiskowe - są to mechanizmy obronne pozwalające przetrwać roślinom w warunkach stresu. Odporność na stres może mieć charakte rkonstytutywny albo indukowany. W pierwszym przypadku mechanizmy obronne występują trwale przez całe życie rośliny. Odporność indukowana to zespół mechanizmów obronnych pojawiających się na skutek działania czynnika stresowego, stresora. Czynniki stresowe środowiska to stresory abiotyczne i biotyczne.
Do czynników abiotycznych zaliczane są: temperatura (wysoka, chłód, mróz), promieniowanie świetlne (wysokie, niskie), susza, niedobór tlenu, czynniki mechaniczne (wiatr, pokrywa śnieżna, pokrywa lodowa), związki chemiczne (zasolenie, toksyny, niedobór minerałów). Do czynników biotycznych zaliczane są: patogeny (grzyby, bakterie, wirusy), rośliny (allelopatia, pasożytnictwo, konkurencja), zwierzęta (zgryzania, pasożytnictwo, deptanie).
Przystosowanie do niekorzystnych warunków środowiska może mieć charakter ADAPTACJI, cech nabytych w drodze ewolucji podlegających zmianie w genomie kolejnych pokoleń oraz AKLIMATYZACJI, cech nabytych w wyniku przebywania osobnika w warunkach stresowych. Aklimatyzacja to modyfikacje niedziedziczone.
ODPOWIEDŹ NA WYMIENIONE WYŻEJ CZYNNIKI OKREŚLANA JEST JAKO:
- A) ODPORNOŚĆ ROŚLIN NA NISKĄ TEMPERATURĘ - zdolność roślin do przetrwania w waurnkach niskich temperatur. Przez niskie temperatury w fizjologii roślin rozumie się zakres 12 stopni Celsjusza do -70 stopni Celsjusza, a nawet niższych.



MECHANIZMY ODPORNOŚCI: Wyróżniane są cztery mechanizmy pozwalające roślinom przetrwać okres mrozów. Pierwszy z nich polega na wykształcaniu izolacji termicznej lub korzystaniu z izolacji zapewnianych przez środowisko. Wierzchołki wzrostu pędu wielu roślin okryte są gęstymi liśćmi. Geofity, a dokładnie ich podziemne części chronione są przed mrozem przez ściółkę i ewentualnie warstwę martwych liści. Opadłe liście zapewniające także izolację podziemnym częściom innych roślin. Rośliny górskie w strefie tropikalnej mogą chronić się przed nocnymi mrozami tworząc rozetę liści zamykającą się nocą. Drugi mechanizm istniejący u roślin polega na obniżaniu temperatury krystalizacji przez zapobieganie zamarzaniu wody w komórce. Przechłodzenie wody jest możliwe dzięki syntezie i gromadzeniu substancji osmotycznie czynnych w wakuoli. Im większe stężenie substancji osmotycznie czynnych tym niższa jest temperatura zamarzania. U roślin drzewiastych rosnących w zimnym klimacie temperatura zamarzania wody w komórce może być obniżona do -50 stopni Celsjusza. Trzeci sposób polega na przemieszczaniu wody do przestrzeni międzytkankowych i zamarzaniu w tych miejscach. Odwodnione tkanki zwiększają stężenie soku komórkowego i obniżają temperaturę zamarzania. Woda może zamarzać w przestrzeni pod okrywą nasienną lub między łuskami pąka. Czwartym mechanizmem pozwalającym przetrwać okres mrozu jest zdolność do krystalizacji wody w przestrzeniach międzykomórkowych. W ścianie komórkowej mogą występować białka AFP (ang anti-freez protein), opóźniające krystalizację i wpływające na kształt powstających kryształów lodu.
 |
Źródło: the-scientist.com - Schemat działania anti-freez proteins.
D) ODPORNOŚĆ ROŚLIN NA ZWIĄZKI TOKSYCZNE - jest to zdolność roślin do przetrwania w środowisku zanieczyszczonym. Najczęściej zanieczyszczenia mają charakter antropogeniczny, mogą to być także związki toksyczne pojawiające się w środowisku naturalnie, w tym związki wytwarzane przez rośliny. Spośród substancji wprowadzanych do środowiska przez człowieka za szkodliwe dla roślin uznawane są dwutlenek siarki (SO2), tlenki azotu (NOx), ozon troposferyczny oraz szereg węglowodorów pojawiających się w powietrzu. Najważniejszą grupą substancji toksycznych pojawiających się w glebach są metale ciężkie, aczkolwiek badany jest także wpływ pestycydów stosowanych w rolnictwie. Wszystkie substancje pochodzenia obcego mogące mieć negatywny wpływ na rośliny określane są jako ksenobiotyki.

SKUTKI OBECNOŚCI METALI CIĘŻKICH W ŚRODOWISKU: większość roślin wykazuje dużą wrażliwość na obecność jonów metali ciężkich w środowisku glebowym. Tylko nieliczne wykształciły mechanizmy pozwalające na przetrwanie w warunkach silnego skażenia. Wniknięcie jonów metali ciężkich do organizmu prowadzi do zaburzenia w działaniu błon, zahamowaniu fotosyntezy oraz zaburzeniu mitochondrialnego transportu elektronów. Inaktywacji ulega wiele enzymów takich jak reduktaza azotanowa, dehydrogenaza izocytrynianowa, dehydrogenaza glukozo-6-fosforanowa, dehydrogenaza jabłczanowa, fosfatazy oraz ATPazy.
MECHANIZMY OBRONNE PRZED DZIAŁANIEM TOKSYCZNYM METALI CIĘŻKICH: Obrona przed toksycznym działaniem metali ciężkich polega na ogrniczeniu ich wnikania do korzeni, a jeżeli pierwszy mechanizm okaże się niewystarczający, ograniczeniu ich rozprzestrzeniania w roślinie. Pierwsza grupa mechanizmów obronnych realizowana jest przez wydzielanie eksudatów korzeniowych zawierających kwasy organiczne, monosacharydy, aminokwasy, kwas poligalakturonowy, i śluzy. Pomocne w ograniczaniu wnikania metali ciężkich są również strzępki grzybów mikoryzowych. Wewnątrz organizmu rośliny metale ciężkie są unieruchamiane w pierwszej kolejności w ścianach komórkowych. Ulegają tam związaniu z składnikami ściany, głównie poprzez reakcję z kwasem poligalakturonowym a także związki fenolowe. Ściana komórkowa jednocześnie staje się mniej przepuszczalna dla roztworów wodnych. Kolejną barierą jest plazmolema, która kontrouje przepływ jonów dzięki obecności kanałów jonowych. Jony metali ciężkich, które dostały się do cytoplazmy są w niej kompartmentowane w wakuoli. W usuwaniu szkodliwych jonów z cytoplazmy uczestniczą peptydy nazywane fitochelatynami. Kompleksy polipeptyd - metal są transportowane do wakuoli i w niej gromadzone. Kompartmentacja w wakuoli zachodzi przede wszystkim w tkankach korzeni, części nadziemne mogą być w ten sposób całkowicie wolne od toksycznego wpływu jonów metali lub narażone na niego w mniejszym stopniu. Chociaż mechanizm ten pozwala uchronić metabolizm komórki roślinnej przed toksycznym działaniem jonów metali ciężkich, to skutkiem ubocznym jego działania może być wielokrotnie wyższe stężenie metali ciężkich w roślinie niż w środowisku glebowym. Rośliny takie nie mogą być wykorzystane jako pokarm dla ludzi lub zwierząt. Zdolnośc akumulacji znalazła jednak zastosowanie w technikach oczyszczania gleb. Wielokrotny siew i zbiór roślin kumulujących metale ciężkie pozwala oczyścić skażone gleby. Proces taki nazywamy fitoremediacją.
E) ODPORNOŚĆ ROŚLIN NA CZYNNIKI BIOTYCZNE - ochronność roślin, obrona przed roślinożercami:
43) OLEOZYNY - są to białka występujące w komórkach roślin. W zależności od gatunku mają masę 15-26 kDa. Występują w organellach magazynujących tłuszcze określanych jako oleosomy. Organella te magazynują tłuszcze zapasowe, głównie tracyloglicerole, i otoczone są monowarstwą lipidową w której zakotwiczone są wystające na zewnątrz oleozyny. Połączone z błoną białka stabilizują struktury zapobiegając zlewaniu się kropli tłuszczu. Występowanie oleozyn stwierdzono zarówno u roślin wyższych oraz mszaków. U zielenic nie stwierdzono obecności białka jednak analiza genomu wskazuje na kodowanie białek o podobnej sekwencji. W genomie Arabidopsis thaliana znajduje się 17 genów kodujących oleozyny, 5 z nich jest aktywnych w nasionach, 3 są aktywne zarówno w nasionach, jak i pyłku, a 9 w komórkach tapetum. Cząsteczka oleozyny składa się z trzech domen. Domena N-końcowa jest hydrofilowa lub amfipatyczna. U Arabidopsis składa się z 68 reszt aminokwasowych. Domena centralna składa się z około 72 reszt i jest hydrofobowa. Tak długiej domeny hydrofobowej nie stwierdzono w żadnym innym białku. Domena C-końcowa może mieć różną długość, a jej odcinek składający się z około 33 reszt jest amfipatyczny i przylega do domeny centralnej i lokalizuje się w pobliżu grup fosforanowych i choliny w monowarstwie fosfolipidowej. Pozostała część domeny C-końcowej nie ma znaczenia funkcjonalnego.
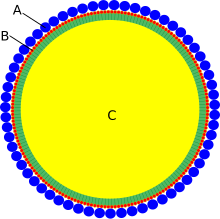
Źródło: Wikipedia. Budowa oleosomu: A - oleozyny, B - monowarstwa fosfolipidowa, C - tłuszcze zapasowe. Oleozyny otaczają organellum zapobiegając łączeniu się kropel tłuszczu.
*Oleosom, sferosom - specyficzne organellum występujące w komórkach roślinnych magazynujące tłuszcze, zwykle triacyloglicerole. Kropla tłuszczu otoczona błoną o połowę cieńszą niż błona komórkowa, składająca się z jednej warstwy białek i fosfolipidów, których końce hydrofilowe skierowane są w stronę cytozolu. Organella te występują w komórkach bielma albo liścieni nasion, a także w komórkach mezokarpu olejowca gwinejskiego oraz owocach awokado. W komórkach tkanek magazynujących tłuszcze może występować do kilkuset tysięcy oleosomów. Występują one również w komórkach mezofilu, gdzie stwierdza się od 1 do 15 sztuk na komórkę oraz w komórkach pyłku i tapetum pylników. Metodami inżynierii genetycznej doprowadzono do powstania oleosomów w komórkach grzybów z gatunku Yarrowia lipolytica. BUDOWA: W komórkach nasion oleosomy to kuliste struktury o średnicy 0,6-2,0 mikrometra. W komórkach mezofilu średnica może wynosić od 1 do 18 mikrometrów. W liściach Sambucus nigra (Bezu czarnego) większe oleosomy stwierdzano w liściach pozostających w cieniu. W mezofilu pszenicy organella zawierały 50-60% tracylogliceroli i 15-40% estrów wosków. Kropla tłuszczu otoczona jest pojedynczą warstwą fosfolipidów oraz białek. Warstwa ta zapobiega zlewaniu się tłuszczu w jedną dużą kroplę. Ważną rolę w stabilizacji struktury oleosomów odgrywają oleozyny. Są to zasadowe białka o masie 15-26 kDa. Część białka zakotwiczona jest w monowarstwie fosfolipidowej a część hydofilowa wystaje poza nią, tworząc zwartą okrywę całego organellum. Zniszczenie części tej okrywy poprzez trawienie trypsyną prowadzi do koalescencji oleosomów. FUNKCJE: Podstawową funkcją oleosomów jest magazynowanie materiałów zapasowych w postaci tłuszczów. Tłuszcze te są zużywane podczas kiełkowania rośliny poprzez włączenie w cykl glioksalanowy. Hyddroliza trriacylogliceroli możliwa jest dzięki obecności w błonie organellum lipaz. W części nasion, na przykład soi i orzecha zimnego, lipaza jest aktywna w glioksysomach, z którymi oleosomy są zasocjowane podczas uruchamiania materiałów zapasowych. Prawdopodobnie także w komórkach mezofilu zmagazynowane tłuszcze stanowią magazyn produktów fotosyntezy. Nie stwierdza się jednak obecności oleosomów w mezofilu traw z anatomią Kranza. Istnieje także hipoteza, według której organella te zapewniają przystosowanie do niskich temperatur. W komórkach mezofilu Diapensia lapponica stwierdzano obecność tylko jednej dużej kroplki tłuszczu w cieplejszych miesiącach i wielu małych kropli w mies^ącach zimnych. BIOGENEZA: Organella powstają w obrębie siateczki śródplazmatycznej. Do błon tej siateczki wprowadzane są oleozyny, co umożliwia rozciąganie warstwy granicznej. Lipidy gromadzone są w cytozolu i ulegają poołączeniu z rejonami siateczki śródplazmatycznej wzbogaconej w stabilizujące białka.
44) Plastyczność fenotypowa - jest to zdolność jednego genotypu do wytworzenia kilku alternatywnych fenotypów. O fenotypie wspólnie z genotypem decydują czynniki środowiskowe. Zmiana warunków środowiska może także prowadzić do zmiany fenotypu. Plastyczność fenotypowa jest jednym z podstawowych mechanizmów adaptacji i obejmuje szereg zmian morfologicznych, fizjologicznych i behawioralnych, obserwowanych u wielu organizmów. Zjawisko jest przedmiotem badań wielu dziedzin biologii, w tym genetyki, genomiki, biologii ewolucyjnej, ekologii, fizjologii i biologii rozwoju. Jest szeroko rozpowszechnione w przyrodzie i w różny sposób może wpływać na tempo ewolucji, przyspieszając ją, spowalniając lub nie zmieniając tempa zmian ewolucyjnych. Zjawisko to jest obserwowane u wielu grup organizmów, jednak ma szczególne znaczenie dla roślin, które są pozbawione możliwości przemieszczania się, a tym samym zmiany warunków życia. Rośliny dostosowują swoją morfologię, anatomię i fizjologię do warunków świetylnych i przez odpowiednią budowę zewnętrzną i wewnętrzną liści, mogą wykazywać zjawisko heterofilii w zależności od warunków środowiska, przejawiają klejstogamię (szczególny rodzaj samozapylenia), wykazują zróżnicowane reakcje obronne w obecności roślinożerców, są zdolne do ruchów oraz konkurencji. Wszystkie te zjawiska są przykładem plastyczności fenotypowej. Zmiany morfologiczne i fizjologiczne mogą także następować w warunkach niskiego stężenia składników pokarmowych. Badania na Thellungiella halophila wykazały, że transporter białkowy ThHAK5 obecny jest w komórkach roślin rosnących w warunkach niedoboru jonów K+. Mesembryanthemum crystallinum, gatunek rośliny rosnący w ekosystemach z niedoborem wody, może w zależności od warunków środowiska przeprowadzać fotosyntezę C3 lub, gdy rośnie w miejscu suchym albo zasolonym, fotosyntezę CAM.
45) Plastydowa oksydaza końcowa, PTOX - jest to enzym występujący w plastydach roślin oraz niektórych glonów. Ortologi (homologi białka lub fragmenty DNA/RNA o wspólnym pochodzeniu ewolucyjnym, których powstanie / rozdział nastąpiło w wyniku specjacji, co odróżnia je od paralogów) wykryto również u sinic. Enzym katalizuje utlenienie plastochinolu z udziałem cząsteczki tlenu. Produktem reakcji jest woda. Reakcja jest alternatywą dla cyklicznego transportu elektronów z udziałem fotoukładu I (PS I) lub fotoukładu II (PS II). Chociaż fizjologiczna rola enzymu nie jest w pełni wyjaśniona, alternatywna droga transportu elektronów z udziałem PTOX uruchamiana jest prawdopodobnie, gdy możliwości akceptorowe PS I są ograniczone. Oksydaza jest również zaangażowana w szlak przenoszenia elektronów określany jako chlorooddychanie. Istnienie tego akceptora elektronów może zapobiegać fotoinhibicji PS I. Enzym jest także niezbędny do przeprowadzenia syntezy karotenoidów. Wszystkie trzy funkcje wiążą się ze zdolnością roślin do radzenia sobie z warunkami stresowymi. Doświadczenia potwierdziły wzrost aktywności oksydazy w warunkach stresowych. Białko wykazuje duże podobieństwo do obecnej w mitochondriach oksydazy alternatywnej (AOX).
U roślin wyższych oraz sinic oksydaza kodowana jest przez jeden gen. U niektórych glonów stwierdzono obecność dwóch genów PTOX1 i pTOX2. Istnienie PTOX było postylowane po raz pierwszy w roku 1982, jednak bezpośrednie dowody zostały przedstawione dopiero w kolejnych latach.
BUDOWA: Plastydowa oksydaza końcowa jest ingegralnym białkiem błonowym. Masa białka została wyznaczona na około 43 kDa u Arabidopsis thaliana. Brak domeny D, która w przypadku oksydazy alternatywnej AOX odpowiada za powstawanie dimeru wskazuje, że funkcjonalnym enzymem jest monomer. Centrum aktywne enzymu znajduje się po stromalnej stronie błony tylakoidów. Niemal wszystkie badane enzymy pTOX zawierały 16-aminokwasową domenę w pobliżu końca C. Jest ona wysoce konserwatywna, a zarazem nie występuje w AOX. Delecja eksonu 8 pokrywającego się z tą domeną wydaje się prowadzić do utraty aktywności. Zarazem wykazano, że białko oksydazy alternatywnej kodowane przez gen AOX2, dzięki obecności właściwego transpeptydu może być importowane do chloroplastów i funkcjonalnie zastępować PTOX. Ważnych dla aktywności enzymu jest 14 reszt aminokwasowych zgrupowanych w trzy klasy: (I) Ala-139, Pro-142, Glu-171, Asn-174, Leu-179, Pro-216, Ala-230, Asp-287, Arg-293 nie są konieczne dla aktywności, (II) Tyr-234 i Asp-295 są niezbędne dla aktywności, (III) Leu-135, His 151 i Tyr-212 są ważne, lecz nie niezbędne dla aktywności (numeracja odnosi się do enzymu z Arabidopsis thaliana). Większość z wymienionych reszt znajduje się w pobliżu sześciu miejsc wiążących dwa atomy żelazy.
EWOLUCJA: Enzym został wykryty u organizmów zdolnych do przeprowadzania oksygenicznej fotosyntezy, czyli u roślin, glonów i sinic. Wysoki stopień podobieństwa między oksydazą alternatywną a plastydową oksydazą końcową wskazuje, że oba enzymy pochodzą od wspólnego przodka, białka zawierającego dwa atomy żelaza. Prawdopodobnie początkowo w czasie transformacji światła beztlenowego na tlenowy, reduktaza tlenu pozwalała pozbyć się tlenu. PTOX pierwotnie powstała u sinic, a AOX u proteobakterii. Oba enzymy pojawiły się u eukariontów w wyniku endosymbiozy, a ich geny były pionowo przekazywane w ewolucji roślin i glonów. Podobieństwo sekwencji genów u glonów i roślin jest wyższe niż 25%. Wysoka konserwatywność wskazuje na powstanie PTOX i AOX jeszcze przed endosymbiozą i braku istotnych zmian od tego punktu. Znane są cyjanofagi zawierające kopie genu kodującego PTOX. Mogą one spełniać rolę wektorów wirusowych przenoszących gen pomiędzy różnymi gatunkami sinic. Uzyskane dane wskazują na wykorzystanie genu przez cyjanofaga w celu zmiany metabolizmu komórki sinicy, tak, aby wytwarzała więcej ATP potrzebnego do syntezy wirusa, a mniej NADPH.
46) POOŚWIETLENIOWY WYRZUT DWUTLENKU WĘGLA PIB (z ang. Post-Illumination Burst of Carbon Dioxide) - jest to zjawisko występujące u części roślin i glonów, polegające na krótkookresowym, intensywnym wydzielaniu ditlenku węgla CO2 w początkowym okresie ciemności przez liście roślin lub glony. ZWIĄZEK POOŚWIETLENIOWEGO WYRZUTU CO2 Z FOTOODDYCHANIEM: Pooświetleniowy wyrzut CO2 został opisany po raz pierwszy w roku 1955 przez J.P. Deckera i od tego czasu zaczęto badać jego zależność od szeregu czynników. Według Deckera (1955) PIB CO2 u roślin C3 jest krótkotrwałą kontynuacją procesu fotooddychania w ciemności, wynikającą z wcześniejszego "wyhamowania" fotosyntezy.. Podstawowym argumentem za fotooddechowym pochodzeniem PIB-u CO2 jest jego brak w atmosferze niskiego stężenia tlenu (1-2%), które ogranicza fotooddychanie. Zauważono, że PIB rośnie wraz ze wzrostem natężenia światła lub wzrostem temperatury. Nawet wzrost temperatury hamujący fotosyntezę, nadal zwiększał wielkość wyrzutu CO2. W dalszych badaniach obserwowano dwa wyrzuty CO2. Pierwszy zależny był wprost proporcjonalnie od natężenia światła poprzedzającego wyrzut i trwał dużo krócej od drugiego. Drugi wyrzut był wielokrotnie dłuższy i mniejszy od pierwszego. Jego wielkość nie zależała od natężenia światła poprzedzającego okres ciemności. Związek wielkości PIB-u z natężeniem fotosyntezy w okresie oświetlania doprowadził do stwierdzenia, iż wyrzut CO2 związany jest z substratami powstającymi podczas fotosyntezy. Szybko dostrzeżono również związek pomiędzy wielkością PIB-u a stężeniem tlenu. Pooświetleniowy wyrzut CO2 był całkkowicie niewidoczny w stężeniu O2 1% i o wiele większy w stężeniu 100% O2 niż przy 21% O2. Jednocześnie obserwowano wyższe natężenie fotosyntezy netto przy stężeniu tlenu 1% oraz brak wpływu stężenia O2 na oddychanie ciemniowe (oddychanie komórkowe). Przy stężeniu O2 1% nie obserwowano obniżania się fotosyntezy wraz ze wzrostem temperatury od 20 do 30 stopni Celsjusza. Badania nad zależnościami pomiędzy PIB-em, fotosyntezą oraz oddychaniem, na świetle i w okresie po oświetlaniu, doprowadziły do odkrycia zjawiska fotooddychania oraz wniosku, iż fotooddychanie jest przez krótki okres kontynuowane w ciemności, czego efektem jest występowanie PIB-u. Zjawisko wyrzutu CO2 obserwuje się również po gwałtownym obniżeniu natężenia światła i nazywane jest międzyoświetleniowym wyrzutem CO2 (PLIB-em) lub po gwałtownym obniżeniu temperatury (postermalny wyrzut CO2 - PLTB). Podobnie jak PIB wyrzut CO2 po obniżeniu natężenia światła jest zależny od stężenia tlenu. Uważa się, że także to zjawisko jest efektem opóźnionego hamowania fotooddychania w stosunku do fotosyntezy. POOŚWIETLENIOWY WYRZUT CO2 U GLONÓW: Zjawisko pooświetleniowego wyrzutu CO2 występuje także u glonów. U Chlamydomonos reinhardii rejestrowano bardzo szybkie pobrania porcji CO2 po włączeniu światła (ang. gulp) oraz bardzo szybkie wydzielenie porcji CO2 po zaciemnieniu. Niewielkie włączanie C14 w związki organiczne i magazynowanie głównie jako węgiel nieorganiczny wskazuje na mały udział procesów fotosyntezy w zjawiskach pobrania i wydzielenia CO2 u Chlamydomonas reinhardii, w relacji światło-ciemność. Gwałtowne zmiany zawartości C14 obserwowano jedynie w puli węgla nieorganicznego. Jego zawartość w komórkach glonów zwiększała się na świetle z 0,5 mM do 2-4 mM i szybko wracała do poziomu z ciemności po zgaszeniu światła. Pooświetleniowy wyrzut CO2 u glonów jest więc związany ze zdolnością do kumulacji nieorganicznych związków węgla na świetle. Na pojawienie się PIB-u u tej grupy organizmów fotooddychanie ma niewielki wpływ. Za niefotooddechową genezą PIB-u u glonów przemawia także fakt braku zmian w wydzielaniu i pobieraniu O2 przy przejściu ze światła do ciemności i odwrotnie. U glonów nie obserwuje się znaczącego wpływu na wielkość PIB-u warunków podwyższających fotooddychanie - obniżonego stężenia CO2, ani też wpływu niskiego (2%) stężenia O2, które hamuje procesy fotooddechowe. Glony prawdopodobnie potrafią sprawnie wiązać dodatkowy CO2 pochodzący z fotooddychania. Akumulacja nieorganicznych postaci węgla w komórkach glonów wymaga nakładów energii i zależy od natężenia światła. Wielkość porcji CO2 pobranej na początku oświetlenia i wydzielonej po zaciemnieniu zależała od gęstości strumienia fotonów jakim były oświetlane komórki Chlamydomonas reinhardii. Badania te przeprowadzono na mutancie, który ma zdolność do zwiększonej akumulacji CO2 wewnątrz komórki, a zgromadzony CO2 był niedostępny dla procesów fotosyntezy (mutant o obniżonej aktywności anhydrazy węglanowej). Zastosowanie inhibitora anhydrazy węglanowej również powodowało wzrost akumulowanego wewnątrz komórki CO2 przy jednoczesnym hamowaniu włączania CO2 w procesie fotosyntezy.
47) PRAWO MITSCHERLICHA, PRAWO PRZYROSTÓW MNIEJ NIŻ PROPORCJONALNYCH, PRAWO OPŁACALNOŚCI NAWOŻENIA - jest to prawo sformułowane przez Eilharda Mitscherlicha w roku 1909, zgodnie z którym wzrost plonów jest tym mniejszy im bardziej ilość dostarczanych składników mineralnych przewyższa optimum fizjologiczne, czyli wprowadzanie do gleby coraz większych dawek nawozów powoduje coraz mniejszy wzrost plonów. FORMUŁA MATEMATYCZNA PRAWA: dy/dx = y = b(a-y) gdzie: x - ilość nawozu, y - plon, a - maksymalna wielkość plonu, b - współczynnik proporcjonalności. 48) PRODUKTYWNOŚĆ ROŚLIN - jest to ilość suchej masy wytworzonej przez roślinę w okresie doby, okresie wegetacji lub w ciągu roku. Termin może być stosowany dla pojedynczej rośliny lub całego łanu. Produktywność roślin jest zależna od intensywności fotosyntezy i oddychania. Produkcja suchej masy zależy od natężenia światła i ilości energii jaka dociera do rośliny w ciągu dnia lub całego okresu wegetacji. Energia światła zamieniana jest z określoną efektywnością na energię chemiczną zgromadzoną w produktach fotosyntezy. Wykorzystanie energii światła przez rośliny obrazuje współczynnik wykorzystania energii świetlnej. Zachodzący jednocześnie proces oddychania prowadzi do zmniejszenia ilości gromadzonej suchej masy. Dla całych zespołów roślinnych określana jest produktywność ekosystemów lub produktywność agrocenoz. W agrocenozach produktywność roślin determinuje plon rolniczy. Proporcja między plonem rolniczym a masą całej rośliny (plonem biologicznym) nazywana jest współczynnikiem plonowania (HI). Ocenę produktywności roślin oraz wpływ czynników endogennych i egzogennych przeprowadza się z użyciem szeregu wskaźników produktywności. Stosowane wskaźniki to:

49) Prohibityny - są to związki chemiczne wytwarzane przez rośliny działające toksycznie na pasożyty. Niejednorodna grupa substancji, wśród których są związki fenolowe, alkaloidy i terpenoidy. Wykazują zdolność do hamowania kiełkowania wzrostu zarodników grzybów, ograniczają wzrost strzępek grzybów, hamują rozwój bakterii oraz namnażanie się wirusów. Właściwości prohibityn wykazują terpenoidy obecne w olejkach eterycznych wytwarzanych przez drzewa iglaste. Podobną rolę odgrywają alkaloidy i flawonoidy wytwarzane przez rosliny motylkowate. Działanie prohibityn może obejmować nie tylko organizm gospodarza, lecz także środowisko wokół niego. Związki syntetyzowane są w odpowiedzi na atak patogenu w wyniku odebrania sygnału hormonalnego.
50) Sieci mikoryzowe (ang. mycorrhizal networks, CMN - common mycorrhizal networks) - sieć pośrednich połączeń pomiędzy roślinami z udziałem grzybów mikoryzowych. Połączenia takie występują we wszystkich ważniejszych ekosystemach lądowych. Strzępki grzybni umożliwiają wymianę szeregu substancji chemicznych pomiędzy elementami sieci. Przenoszone mogą być: związki węgla, sole mineralne, woda, substancje sygnałowe oraz allelopatiny. Sieci mikoryzowe łączą rośliny autotroficzne, myko-heterotrofy oraz częściowe myko-heterotrofy. Połączenie różnorodnych organizmów przez strzępki grzybów ułatwia im przetrwanie i wzrost, zapewniając różnorodnej gatunkowo grupie wspólną stabilność w zmieniających się warunkach środowiska. Około 75% badanych gatunków roślin korzysta z sieci mikoryzowych.
SUBSTANCJE PRZENOSZONE W SIECI MIKORYZOWEJ: Liczne badania wykazały możliwość transportowania pomiędzy roślinami przez sieci mikoryzowe związków węgla, fosforu, azotu, wody, związków chemicznych zapewniających obronę i allelopatin. Sieci mikoryzowe umożliwiają transport substancji mineralnych z miejsc zasobnych do roślin rosnących w mniej korzystnych warunkach. Zarazem możliwe jest przeniesienie związków węgla z roślin rosnących w warunkach dobrego oświetlenia do roślin zacienionych. Do odwzorowania procesów zachodzących z udziałem strzępek grzybów przydatny jest model donor - akceptor (ang. model source - sink). Dobrze rozwinięta sieć zwiększa prawdopodobieństwo zakażenia grzybem mikoryzowym kolejnych roślin. Wykazano także zwiększony sukces reprodukcyjny roślin uczestniczących w wymianie substancji w sieciach mikoryzowych.
RODZAJE SIECI MIKORYZOWYCH: Sieci mikoryzowe mogą powstawać w wyniku mikoryzy arbuskularnej oraz ektomikoryzy. Sieci mikoryzowe arbuskularne tworzone są głównie z udziałem grzybów z gromady Glomeromycota. Tego rodzaju połączenia mikoryzowe są domunującym typem współpracy grzybów oraz roślin. Dotychczas poznano około 150-200 gatunków grzybów tworzących ten typ mikoryzy, jednak może być ich znacznie więcej. Charakterystyczną cechą tej mikoryzy jest niska specyficzność wobec gospodarza, chociaż zwykle gatunki grzybów mają określone preferencje względem gatunku rośliny. Sieci oparte o ektomikoryzę składają się z roślin tworzących mikoryzę ze ściśle określonymi gatunkami grzybów. Znanych jest około 10 tysięcy gatunków grzybów wchodzących w tego rodzaju relacje z roślinami. Gatunki te należą do wielu grup systematycznych. Sieci ektomikoryzowe dominują w lasach strefy umiarkowanej oraz lasach borealnych, obejmując raczej konkretne gatunki.
KORZYŚCI Z POWSTANIA SIECI: Efektem wykształcenia sieci mikoryzowych może być transfer substancji pomiędzy miejscami o różnej ich dostępności. Umożliwia to kolonizację powierzchni, na których jeden ze składników, występujący w niedostatecznej ilości, ograniczałby wzrost roślin. Korzyści rozpatrywane są także w zakresie zwiększenia możliwości rozwoju roślin i grzybów mikoryzowych. Wzrost liczebności roślin pozwala rozwijać się grzybom, i odwrotnie. Doświadczenia wskazują na znaczący wpływ sieci ektomikoryzowych na rozwój siewek drzew iglastych. Przepływ węgla organicznego między roślinami a ich partnerem grzybowym podlega kontroli. Rośliny mają możliwość ograniczenia przepływu węglowodanów tak, aby uniknąć pasożytnictwa grzybów. Kontrola może polegać także na nietworzeniu połączeń z grzybami w warunkach dobrego zaopatrzenia w składniki mineralne. Z kolei w warunkach niedoboru substancji mineralnych rośliny zwiększają ilości wydzielanych do gleby strigolaktonów - substancji stymulujących rozwój grzybów mikoryzowych. Postuluje się, że współpraca mikoryzowa jest promowana, gdy u partnerów występują nadwyżki zasobów. Z tego względu sieci mikoryzowe mogą być rozpatrywane jako platforma handlowa, w której wielu partnerów dokonuje inwestycji, jednak nie są one jednakowo opłacalne dla wszystkich. Badania sieci, w których len i sorgo połączone były komponentem grzybowym w postaci Glomus mosseae lub G. intraradices pokazały, że inwestycji w postaci związków organicznych dokonują głównie rośliny sorga (70% związków organicznych w sieci z udziałem G. intraradices i 71% w sieci z G. mosseae). Korzyści w postaci związków azotu i fosforu odnosiły w sieci z G. intraradices głównie rośliny lnu (98% P 80% N), a w sieci z G. mosseae korzyści dzielone były stosunkowo równo (48% P i 52% N dla lnu, t2% P i 48% N dla sorga). Sieci mikoryzowe nie są więc formą "systemu socjalistycznego", w którym bogactwa są dzielone "po równo", lecz przypominają "system kapitalistyczny", w którym każdy organizm stara się osiągnąć maksymalny zysk.
Poza możliwością przekazywania substancji pokarmowych ważnym efektem istnienia sieci mikoryzowych jest wspólna obrona przed roślinożercami. W doświadczeniach laboratoryjnych wykazano, że zgryzanie przez gąsienice Spodoptera litura liści pomidora indukowało reakcje obronne u sąsiadujących roślin połączonych strzępkami Funneliformis mosseae. Substancją sygnałową przenoszoną przez grzyba był prawdopodobnie kwas jasmonowy. Stałe wytwarzanie substancji obronnych jest kosztowne, dlatego synteza wielu z nich jest indukowana uszkodzeniami powstającymi podczas ataku. Przekazywanie sygnału pozwala uruchomić reakcje obronne bez ponoszenia strat przez każdą z roślin. Około 10-15% gatunków roślin naczyniowych nie wchodzi w interakcje z grzybami mikoryzowymi. Jednym z takich gatunków jest Arabidopsis thaliana wykorzystywany jako roślina modelowa.
SIECI MIKORYZOWE, A MYKO-HETEROTROFY: Znanych jest około 400 gatunków roślin zdolnych do pobierania związków organicznych z grzybów. U niektórych z tych roślin zaobserwowano miksotrofię, pobieranie związków organicznych z grzybów lub przeprowadzanie fotosyntezy, w zależności od dostępu do światła. Badania na myko-heterotrofach, grzybach mikoryzowych oraz roślinach zielonych tworzących sieci mikoryzowe w lasach tropikalnych wykazują, że węgiel i azot u wszystkich trzech grup organizmów pochodzą z tych samych źródeł, zawartość azotu jest jednak wyższa w grzybach niż w roślinach. Bez wątpienia pozbawione chlorofilu myko-heterotrofy korzystają z węgla organicznego dostarczonego przez grzyby mikoryzowe. Ze względu na istnienie sieci mikoryzowych związki organiczne mogą pochodzić od innych roślin połączonych z grzybami. Ekologiczna rola myko-heterotrofów jako pasożyta nie jest jednak oczywista. Stwierdzono, że Sarcodes sanguinea otrzymuje związki organiczne od Rhizopogon ellenae, pozostającego jednocześnie w mikoryzie z Abies magnifica (Jodłą wspaniałą), zarazem w pobliżu miejsc, gdzie rośnie S. sanguinea korzenie A. magnifica były lepiej rozwinięte. Także liczebność R. ellenae maleje wraz ze wzrostem odległości od S. sanguinea. Nie jest jasne, czy myko-heterotrof stymuluje wzrost swojego pośredniego żywiciela, czy też relacja ma charakter mutualistyczny.
ROLA SIECI MIKORYZOWYCH W EKOSYSTEMACH: Połączenie siecią mikoryzową może powodować dodatnie sprzężenie zwrotne pomiędzy osobnikami dorosłymi a siewkami roślin jednego gatunku i w efekcie prowadzić do dominacji tego gatunku w ekosystemie. Wykazano, że możliwe jest uzyskanie przewagi prowadzącej do dominacji jednego gatunku drzewa w drzewostanie. W przypadku Dicymbe corymbosa, drzewa występującego w Gujanie, gdy sadzonki mają dostęp do sieci mikoryzowej charakteryzują się wyższą przeżywalnością, zwiększoną liczbą liści oraz są wyższe niż sadzonki bez dostępu do sieci. Wśród przebadanych gatunków u 48% z nich siewki uzyskiwały korzyści z istnienia sieci mikoryzowych. Dla 27% istnienie sieci było neutralne, a u 25% obserwowano negatywne skutki przyłączenia. Siewki odnoszą korzyści głównie w sieciach ektomikoryzowych (75% badanych gatunków), które są bardziej specyficzne gatunkowo. W sieciach mikoryzowych arbuskularnych tylko siewki 42% gatunków uzyskiwały korzyści.
Dysproporcje między ilością inwestowanych związków organicznych a otrzymywanych od komponentu grzybowego związków mineralnych mogą być wykorzystane w agroekosystemach do zwiększenia plonu gatunku odnoszącego większe korzyści z istnienia sieci mikoryzowej.
51) Sok mleczny, lateks naturalny - jest to nieprzezroczysta zawiesina wodna gromadzona w wakuolach komórek mlecznych roślin. Skład chemiczny, wielkości cząstek zawiesiny i jej barwa oraz wydzielane ilości soku mlecznego są zróżnicowane w zależności od gatunku rośliny. W skład zawiesiny wchodzić mogą z różnym udziałem: węglowodany (w tym skrobia), żywice, woski, gumy, białka, olejki lotne, alkaloidy, garbniki, kauczuk i inne związki chemiczne, udział wody wynosi 50-80%. Sok mleczny najczęściej ma barwę białą, bywa także żółty i pomarańczowy. Obecność lub brak soku mlecznego i jego barwa wykorzystywana jest jako cecha taksonomiczna. Sok mleczny posiadają m.in. przedstawiciele makowatych, wilczomleczowatych i astrowatych. Sokiem mlecznym nazywany jest także biały lub innej barwy płyn wydzielający się z uszkodzonych owocników niektórych gatunków grzybów, tzw. mleczko.

Źródło: Wikipedia. Wyciekający sok mleczny z przełamanej łodygi glistnika jaskółcze ziele.
ZASTOSOWANIE: Sok mleczny niektórych roślin ma znaczenie użytkowe - może służyć do wyrobu kauczuku naturalnego, gutaperki oraz produktów leczniczych lub narkotycznych (np. lactucarium, opium). Głównym gatukiem dostarczającym surowca do produkcji kauczuku naturalnego jest kauczukowiec brazylijski, którego sok mleczny zawiera od 20 do 60% kauczuku. Inne gatunki użytkowe do wytwarzania kauczuku to pozostali przedstawiciele rodzaju kauczukowiec (Hevea), poza tym figowce (Ficus) oraz mniszek kok-sagiz (Teraxacum kok-saghyz). W sumie znanych jest ok. 200 gatunków roślin kauczukodajnych. W celu produkcji kauczuku naturalnego sok mleczny stabilizowany jest substancjami alkalicznymi np. amoniakiem. Ostateczny produkt w postaci kauczuku naturalnego stanowiącego surowiec w przemyśle gumowym uzyskuje się w wyniku kagulacji i zagęszczenia. Zagęszczenie zawartości kauczuku wykonywane jest zwykle poprzez odwirowanie lub częściowe odparowanie wody. Koagulacja następuje po zakwaszeniu, zamrożeniu lub dodaniu soli rozpuszczalnych w wodzie.
*Kauczuk naturalny, cis-1,4-poliizopren - jest to polimer pochodzenia naturalnego należący do politerpenów. Cząsteczki polimeru mogą składać się z 320 do 35 000 reszt izoprenowych, co odpowiada masie cząsteczkowej od 100 tysięcy do kilku milionów Da. W cząsteczkach kauczuku występują wyłącznie wiązania podwójne w konfiguracji cis. Z kauczuku wytwarzanych jest ponad 40 000 wyrobów w tym ponad 400 wyrobów medycznych. Substratami w biosyntezie kauczuku są kwas mewalonowy i pirofosforan izopentylu. Enzymy niezbędne do przeprowadzenia reakcji obecne są w soku mlecznym a energia pochodzi z glikolizy, której zachodzenie zaobserwowano w wydzielinie. PRODUKCJA PRZEMYSŁOWA: Wykorzystanie kauczuku przez człowieka rozpoczęło się w Ameryce Centralnej jeszcze przed jej odkryciem przez Kolumba. Stosowaną nazwę uzyskiwanej z soku drzew elastycznej masie nadali Indianie. Europejczycy przybyli na kontynent amerykański nie tylko przejęli technikę pozyskiwania kauczuku lecz także rozprzestrzenili uprawy na inne obszary. Kauczuk pozyskiwany jest głównie z drzewa kauczukowca brazylijskiego (Hevea brasiliensis), uprawianego w wilgotnym, tropikalnym klimacie. Jeden hektar upraw to od 350 do 500 drzew. Z każdego drzewa można uzyskać około 5 kg suchego surowca, co odpowiada rocznej wydajności upraw na poziomie 2000 kg/ha. Kauczuk pozyskiwany jest z drzew w wieku pomiędzy 5 a 20-25 lat. Rano wykonuje się na drzewach nacięcia w kształcie litery V. Poniżej nacięć umieszczane jest naczynie, do którego spływa sok mleczny. Pozyskany lateks zbierany jest po 5-8 godzinach od nacinania drzew. Zebrany płyn może być oczyszczany i stabilizowany a następnie przewożony do zakładów przemysłowych wytwarzających przedmioty z kauczuku lub już na plantacji suszony i formowany w bryły o masie 33 kg wykorzystywane w przemyśle. Wysokiej jakości kauczuk ma jednolitą konsystencję i żółtą barwę. Kauczuk rozpuszcza się w węglowodorach alifatycznych i aromatycznych oraz w węglowodorach chlorowanych. Roztwory kauczuku naturalnego zawierające substancje wulkanizujące stosuje się jako kleje.

Źródło: Wikipedia. Wzór strukturalny kauczuku naturalnego.
*Gutaperka - jest to substancja pochodzenia naturalnego, pozyskiwana i wykorzystywana w podobny sposób jak kauczuk naturalny, z tą różnicą, że gutaperka jest mniej elastyczna.

Źródło: Wikipedia. Struktura chemiczna gutaperki.
Cząsteczki gutaperki składają się z reszt C5, których liczba dochodzi do kilkuset. W polimerze występują wiązania podwójne wyłącznie w konfiguracji trans. Niektórzy autorzy polimer określają nazwą gutta (lub guta), zaś gutaperką mieszaninę takiego polimeru z alkoholowymi pochodnymi triterpenów. Mieszaniny takie są naturalnym składnikiem soku mlecznego niektórych roślin. Nazwa gutaperka pochodzi z języka malajskiego: getah = guma, pertja = drzewo z którego pozyskuje się substancję. WYSTĘPOWANIE: Gutaperka występuje w soku mlecznym wielu gatunków roślin. Największe znaczenie ma gutaperkowiec (Palaquium gutta) oraz zwięźla kulista (Mimusops balata). Gutaperkowiec jest drzewem rosnącym na Półwyspie Malajskim i w Indonezji. Występuje także w dużych ilościach u eukomii wiązowatej (Eucommia ulmoides). Spośród gatunków europejskich stwierdzona została w dużych ilościach w trzmielinie brodawkowatej i zwyczajnej. WŁAŚCIWOŚCI: Wyróżniane są dwie formy gutaperki: alfa i beta. Formy te przechodzą w postać amorficzną i stają się płynne w różnym zakresie temperatur mieszczącym się pomiędzy 40 a 70 stopni Celsjusza. Dla formy beta temperatura topnienia jest niższa niż dla formy alfa. Forma alfa jest charakterystyczna dla gutaperki wytwarzanej przez drzewa. Obie formy zawierają wiązania trans, a różnią się wielkością cząstek. Gęstość 0,95 - 1,02 g/cm3. Jest mniej elastyczna niż kauczuk, wykazuje jednak większą odporność chemiczną oraz dużą odporność na czynniki klimatyczne. Ulega starzeniu od tlenu atmosferycznego, przekształcając się w kruchą masę, ale można temu zapobiec odpowiednimi dodatkami. Rozpuszczalna w węglowodorach aromatycznych i chlorowanych, ulega kwasom tlenowym. Jest dobrym izolatorem elektrycznym.

52) Starzenie się roślin - jest to końcowy etap rozwoju rośliny, charakteryzujący się osłabieniem procesów życiowych. Ostatecznym rezultatem procesu starzenia jest śmierć. Starzenie się może obejmować całą roślinę lub poszczególne jej organy. Starzenie się całego organizmu roślinnego zachodzi po wytworzeniu kwiatów i owoców u roślin monokarpicznych. U bylin na koniec sezonu wegetacyjnego starzeją się i obumierają części nadziemne. U roślin polikarpicznych występuje zjawisko starzenia się niektórych organów. Starzeniu podlegają kwiaty, owoce i liście. ZMIANY W OBRĘBIE KOMÓRKI: o rozpoczęciu procesu starzenia decydują zarówno czynniki genetyczne, jak i środowiskowe. W początkowym etapie dochodzi do spadku zawartości auksyn, cytokinin i giberelin w starzejącej się tkance, jednocześnie wzrasta stężenie etylenu i kwasu abscysynowego. Widocznym objawem starzenia jest rozpad chlorofilu i utrata turgoru komórek. Degradacji ulega ultrastruktura chloroplastów. Zanikają błony lamelarne, zanikają rybosomy, zwiększa się ilość i rozmiar globul plastydowych. Zmniejsza się także ilość rybosomów w cytozolu, dochodzi do rozpadu retikulum endoplazmatycznego, aparatów Golgiego i tonoplastu. W końcowym etapie zniszczeniu ulega ultrastruktura jądra komórkowego. Zmiany hormonalne prowadzą do syntezy de novo licznych enzymów hydrolitycznych takich jak proteazy, nukleazy, lipazy, amylazy, pektynazy i celulazy. Powstające w reakcjach katalizowanych przez te enzymy związki proste są odprowadzane z tkanek starzejących się do innych części rośliny. ODCINANIE ORGANÓW: W strefie klimatu umiarkowanego jesienią dochodzi do zrzucania liści. Po odprowadzeniu przydatnych dla organizmu związków ze starzejącego się organu dochodzi do wytworzenia warstwy komó®ek nazywanej strefą odcinającą. Komórki tej strefy w wyniku działania etylenu syntetyzują enzymy hydrolizująe polischarydy blaszki środkowej. Osłabienie połączeń pomiędzy komórkami prowadzi do oderwania się organu. Warstwa odcinająca tworzy się u nasady ogonka liściowego. Poza liśćmi strefa odcinająca wytwarzana ejst także u nasady szypułki kwiatowej, tworzy się ona jeżeli nie doszło do zapłodnienia lub jest ono niepełne. W przypadku owoców powstawać może jedna strefa odcinająca między szypułką a pędem, tak jest u owoców ziarnkowych lub dwie strefy odcinające, między szypułką a pędem i między szypułką a owocnią, tak jest u owoców pestkowych. Po odpadnięciu organu strefa odcinająca pokrywana jest substancjami chroniącymi przed patogenami takimi jak kutyna, garbniki. Proces tworzenia warstwy odcinającej jest stymulowany przez etylen i kwas absycysynowy. Nazwa tego drugiego związku pochodzi od ang. abscission - odcinanie.

Źródło: Wikipedia. Pierwszy etap starzenia się roślin to rozpad chlorofilu.
53) Symplast - jest to zespół protoplastów połączonych plasmodesmami i otoczony plasmolemmą (błoną komórkową). Plazmodesmy pozwalają na bezpośredni przepływ (dyfuzja) małych cząsteczek między sąsiadującymi komórkami, np. cukry, aminokwasy. Większe cząsteczki mogą być również transportowane za pomocą filamentów aktynowych.
54) Apoplast - jest to zespół ścian komórkowych, wnętrz martwych komórek i przestworów międzykomórkowych w organizmie roślinnym. Połączone ze sobą i wysycone wodę ściany komórkowe razem z wypełnionymi wodą wnętrzami martwych komórek przewodzących są głównymi drogami transportu wody w roślinie. W ścianach komórkowych ruch wody możliwy jest dzięki zjawisku imbibicji. Wypełnione powietrzem przestwory międzykomórkowe stanowią rezerwuary gazów i umożliwiają roślinie prowadzenie wymiany gazowej z otoczeniem.

Źródło: Wikimedia. Apoplast i symplast.
55) SZLAK PENTOZOFOSFORANOWY (SZLAK HEKSOZOMONOFOSFORANOWY, SZLAK FOSFOGLUKONIANOWY) - jest to ciąg reakcji biochemicznych podczas których glukozo-6-fosforan jest utleniany do rybulozo-5-fosforanu oraz wytwarzany jest NADPH. Głównym celem jest dostarczanie komórce NADPH niezbędnego do przeprowadzania reakcji redukcji w cytoplazmie oraz synteza pentoz.
Reakcje szlaku zachodzą w komórkach roślinnych w cytoplazmie oraz plastydach. Zachodzi takżę w komórkach zwierzęcych - przede wszystkim w komórkach tkanki tłuszczowej, gruczołów mlekowych, wątroby, erytrocytów, jąder i kory nadnerczy. Opisane poniżej reakcje nazywane są oksydacyjnym szlakiem pentozofosforanowym. Te same enzymy wykorzystywane są w szlaku określanym jako redukcyjny szlak pentozofosforanowy, służącym do odtworzenia z aldehydu fosfoglicerynowego rybulozo-1,5-bisfosforanu w fazie regeneracyjnej cyklu Calvina, zachodzącej w fotosyntetyzujących komórkach roślinnych. Wykorzystanie tych samych enzymów w cyklach reakcji o różnym efekcie końcowym pokazuje swoistą oszczędność ewolucji. SUMARYCZNA REAKCJA SZLAKU: glukozo-6-fosforan + 2 NADP+ + H20 -> rybozo-5-fosforan + 2 NADPH + 2 H+ + CO2.
PRZEBIEG SZLAKU: W przebiegu szlaku pentozofosforanowego można wyróżnić dwie fazy. Pierwsza - faza oksydacyjna, podczas której powstaje NADPH oraz druga - faza nieoksydacyjna, podczas któ®ej powstają pentozy oraz cukry o 3, 4 i 7 atomach węgla.
FAZA OKSYDACYJNA: Podczas tej fazy glukozo-6-fosforan zostaje przekształcony w rybulozo-5-fosforan. Jednocześnie dwie cząsteczki NADP+ zostają zredukowane do NADPH+H+.
    |
 - Soliród zielny na brzegu słonego jeziora Baskunczak, źródło: Wikipedia.
- Soliród zielny na brzegu słonego jeziora Baskunczak, źródło: Wikipedia.








Reakcje tego etapu są odwracalne. Jeśli zapotrzebowanie w komórce na NADPH jest większe niż na rybozo-5-fosforan, zostaje on przekształcony we fruktozo-6-fosforan i aldehyd 3-fosfoglicerynowy, które są metabolitami glikolizy, zgodnie przedstawionymi reakcjami. Jeśli natomiast zapotrzebowanie na rybozo-5-fosforan jest znacznie większe niż na NADPH, transketolaza i transaldolaza przekształcają fruktozo-6-fosforan i aldehyd 3-fosfoglicerynowy, pobrane z glikolizy, w rybozo-5-fosforan.

SZLAK PENTOZOFOSFORANOWY A MALARIA: Zauważono, że genetycznie uwarunkowany niedobór dehydrogenazy glukozo-6-fosforanowej występuje znacznie częściej u Amerykanów pochodzenia afrykańskiego (11%) niż Amerykanów pochodzących z innych kontynentów. Zostało wykazane, że niedobór enzymu szlaku pentozofosforanowego jest czynnikiem chroniącym przed malarią. Pasożyt - Plasmodium falciparum - wywołujący chorobę do rozwoju potrzebuje glutationu, będącego metabolitem opisanego powyżej szlaku.
56) SZTUCZNE NASIONA - są to zarodki roślin uzyskane metodami biotechnologicznymi w hodowli in vitro, służące do rozmnażania elitarnych odmian roślin Zarodki uzyskiwane są w wyniku przeprowadzenia embriogenezy somatycznej. Po wykształceniu mogą być suszone lub utrwalane w innej formie, co umożliwia ich przechowywanie podobnie jak nasion naturalnych. Technologia uzyskiwania sztucznych nasion rozwijana jest od lat 60. XX wieku, jednak jej zastosowanie komercyjne jest ograniczone. Nasiona uzyskiwane w wyniku embriogenezy somatyznej zapewniają pełną kontrolę nad genotypem uzyskiwanych roślin, co nie jest możliwe przy rozmnażniu płciowym. Rozwój technologii sztucznych nasion jest atrakcyjną perspektywą w rolnictwie oraz sadownictwie i leśnictwie. UZYSKIWANIE: Stosowane są dwie metody pordukcji sztucznych nasion - pierwsza z nich określana jako mokra, polega na otoczeniu zarodka kapsułką z hydrożelu. Otoczka żelowa spełnia funkcję analogiczną do bielma. Zwykle ma skład pożywki MS (Pożywka Murashuge & Skoog) oraz zawiera substancję żelującą, którą może być alginian, karaginian, agar, pektan sodu, guma tragakant lub karboksymetyloceluloza. Stwierdzono, że dodatek sacharozy do otoczki nie wpływa na proces kiełkowania. Otoczka z hydrożelu może być dodatkowo pokryta warstwą chroniącą przed utratą wody. Druga z metoda polega na wysuszeniu zarodka w stadium dojrzałości i pokryciu warstwą zabezpieczającą. Jest to tak zwana metoda sucha. Jedną ze stosowanych do tego celu substancji jest polioksyetylen. W drugiej fazie dojrzewania zarodki przeznaczane na nasiona suche poddawane są działaniu kwasu absysynowego. Zapewnia to odporność zarodków na suszenie. Uzyskane tą metodą sztuczne nasiona mogą być przechowywane przez co najmniej 169 tygodni w temperaturze -25 stopni Celsjusza. Sztuczne nasiona uzyskane metodą suchą są pozbawione materiałów zapasowych, dlatego przed wysianiem poddawane muszą być procesowi regeneracji i hodowli na pożywkach. Dopiero sadzonki z takich roślin wysadzane są do gleby. Uzyskane nasiona muszą wykazywać się zdolnością do kiełkowania i konwersji (rozwoju korzonka zarodkowego i merystemu pędu z wykształceniem właściwego fenotypu).
57) TRANSPORT FLOEMOWY - jest to podstawowy mechanizm transportu asymilatów na duże odległości przez elementy łyka. Daleki transport asymilatów odbywa się w trzech etapach. W pierwszym etapie nazywanym załadunkiem floemu asymilaty przenoszone są z tkanek donora do rurek sitowych. W drugim etapie odbywa się transport właściwy, czyli przepływ asymilatów. Trzeci etap, nazywany rozładowaniem floemu polega na przeniesieniu asymilatów z rurek sitowych do tkanek akceptora. Tkanki, z których pochodzą substancje organiczne transportowane floemem nazywane są donorem. Zwykle są to tkanki liścia, jednak dokadne przyporządkowanie funkcji donora poszczególnym organom jest niemożliwe. W początkowym okresie wzrostu młode liście wymagają dostarczenia związków organicznych niezbędnych do budowy organu. Akceptorami są tkanki odbierające związki przenoszone przez floem. Typowymi akceptorami są tkanki korzenia i kwiaty. Niektóre organy pełnią funkcję organu spichrzowego gromadząc asymilaty w określonej fazie rozwoju rośliny, a stają się donorem na kolejnym etapie.

Źródło: Wikipedia. Obrączkowanie polegające na usunięciu kory wtórnej prowadzi do śmierci drzewa. Asymilaty nie mogą być przeniesione do korzeni.
ZAŁADUNEK FLOEMU: Przekazywanie asymilatów i innych związków do rurek sitowych odbywa się za pośrednictwem komórek miękiszowych. Przeniesienie związków chemicznych z komórek towarzyszących do rurki sitowej odbywa się na drodze transportu aktywnego i przebiega wbrew gradientowi stężeń. Stężenie w rurkach sitowych może być 10-20 razy wyższe niż komórkach miękiszu. Załadunek odbywa się przez symplast, z wyorzystaniem plazmodesm łączących komórki towarzyszące z rurkami sitowymi oraz apoplastem. Przy drugim sposobie załadunku konieczne jest przeniesienie związków przez plazmolemę dwukrotnie, dlatego tą drogą wiąże się ze znacznym zapotrzebowaniem na energię.
SKŁAD SOKU FLOEMU: Sok floemu jest wodnym roztworem, związków chemicznych, głównie organicznych. Do 90% suchej masy soku stanowią cukry. Cechą charakterystyczną jest brak cukrów redukujących. Zjwisko to zostało nazwane "heksofobią". Transportowana jest przede wszystkim sacharoza, a także w zależności od gatunku rafinoza, stachioza, werbaskoza, ajugoza. Drugą grupa składników soku są polialkohole sześciowęglowe: sorbitol i mannitol. W soku znajdują się także związki azotu takie jak aminokwasy i amidy. U roślin motylkowych formą transportową azotu jest kwas alantoinowy i cytrulina wytwarzne w brodawkach korzeniowych. Pozostałe składniki soku to kwasy organiczne, nukleotydy, ortofosforany, ATP, witaminy, fitohormony.
TRANSPORT NA DUŻE ODLEGŁOŚCI: Mechanizm transportu związków w rurkach sitowych nie jest dokładnie wyjaśniony. Powszechnie przyjmuje się sformułowaną przez Ernsta Muncha w roku 1930 hipotezę przepływu masowego. Zgodnie z hipotezą sok floemu przemieszcza się pod wpływem ciśnienia hydrostatycznego, które jest efektem wysokiego stężenia sacharozy w miejscu załadunku i związanego z nim niskiego potencjału wody. W pobliżu akceptora, stężenie sacharozy jest niskie, a potencjał wody wysoki.
ROZŁADUNEK FLOEMU: Rozładunek floemu odbywa się podobnie jak załadunek na drodze transportu aktywnego. Może zachodzić zarówno przez symplast, jak i apoplast. Wszystkie procesy rozładunki wymagają zużycia ATP. Jeżeli akceptorem jest organ spichrzowy rozładunek odbywa się przez apoplast. Na powierzchni ścian komórkowych akceptora lub w cytozolu następuje hydroliza sacharozy i oligosacharydów w wyniku działania inwertazy.
58) Utlenianie fotosensybilizowane, fotooksydacja - jest to proces przeniesienia elektronu (typ I) lub energii (typ II) z cząsteczki barwnika absorbującego (fotosensybilizator) promieniowanie widzialne, ultrafioletowe lub bliską podczerwień, na inną cząsteczkę chemiczną. W typie I dochodzi do utlenienia donora wzbudzony stan trypletowy elektronu przez fotosensybilizatora lub fotosensybilizator przechodzi do stanu wzbudzonego singletowego, jeśli jednak nie odda energii i nie powróci do stanu podstawowego może przejść w bardziej trwały stan trypletowy, co wiąże się z dużym prawdopodobieństwem udziału w reakcji utleniania fotosensybilizowanego. Utlenianie fotosensybilizowane jest przyczyną powstawania w organizmach żywych reaktywnych form tlenu będących przyczyną stresu oksydacyjnego i uszkodzenia związkó∑ chemicznych obecnych w komórkach. W roślinach, posiadających chlorofile mogące spełniać rolę fotosensybilizatora proces fotooksydacji może prowadzić do uszkodzenia błon poprzez utlenienie lipidów chloroplastowych. Aby zapobiec przekazywaniu energii stanu trypletowego na inne cząsteczki rośliny wykształciłī systemy antenowe w których posiadają zarówno cząsteczki chlorofilu jak i karotenoidów. Jeśli stan wzbudzenia chlorofilu nie może być przekazany do centrum reakcji fotoukładu i chlorofil przejdzie w stan trypletowy jego energię przejmują karotenoidy i rozpraszają ją w postaci ciepła co chroni chloroplast przed fotooksydacją.
 |
| Źródło: Wikipedia. Antocyjany w komórkach aleuronowych nadają barwę ziarniakom kukurydzy. |
59) Warstwa aleuronowa, komórki aleuronowe - jest to zewnętrzna warstwa komórek bielma w ziarniakach. Komórki warstwy aleuronowej są prostopadłościanami o regularnych kształtach, a w ich cytozolu zawieszone są liczne ziarna aleuronowe i ziarna skrobi. U kukurydzy w komórkach aleuronowych mogą występować znaczne ilości antocyjanów nadających barwę ziarniakom. W fazie imbibicji, pierwszym etapie kiełkowania w warstwie aleuronowej zachodzi synteza alfa-amylazy. Proces jest indukowany przez gibereliny, któ®e do komórek aleuronowych docierają z zarodka. W zarodku gibereliny powstają w efekcie hydrolizy koniugatów. W warstwie aleuronowej efektem działania giberelin jest aktywacja aparatu biosyntezy białek, pobudzenie transkrypcji genu kodującego alfa-amylazę oraz uruchomienie mechanizmu umożliwiającego transport przez błonę komórkową wytworzonych białek. Wydzielona alfa-amylaza katalizuje reakcję hydrolizy skrobi w bielmie.
60) Współczynnik oddechowy, RQ - jest to stosunek objętości wydalanego CO2 do ilości pobranego tlenu. WO = CO2 wydzielone/ O2 zużyte. Współczynnik jest użyteczny ponieważ objętość zużytego tlenu i wydalonego dwutlenku węgla zależy od rodzaju metabolizowanej substancji. Mierząc WO można określić jakie substancje są źródłem energii danego organizmu. Dla cukrów prostych (glukoza) współczynnik oddechowy wynosi 1, lecz np. dla tłuszczów nasyconych (glicerol trójestrowy) wynosi 0,66. Wielkość współczynnika oddechowego zależeć będzie także od temperatury i od dostępności tlenu. U ssaków w casie embriogenezy współczynnik oddechowy zwiększa się znacznie w stadium wolnej blastocysty.
61) Wpółczynnik plonowania - KLIK.
62) Współczynnik wykorzystania energii świetlnej:

63) Wysmalanie roślin - jest to zjawisko występujące w zimie, kiedy rośliny nie są okryte śniegiem, a ponadto wieją silne wiatry i jest słoneczna pogoda. W takich warunkach rośliny transpirują wodę i jednocześnie nie mogą jej uzupełnić, ponieważ gleba jest zamarznięta, więc więdną, zasychają i giną.
64) Ziarno aleuronowe (gr. aleuron - mąka) - są to w przybliżeniu kuliste, struktury białkowe, otoczone pojedynczą błoną biologiczną, powstające przez odwodnienie i tężenie licznych drobnych wakuoli, rozmieszczonych równomiernie w cytoplazmie. Powstają w końcowym etapie dojrzewania nasion, a zanikają (zostają zużyte) na początku kiełkowania, dostarczając substancji odżywczych kiełkującemu zarodkowi. BUDOWA: Ziarna aleuronowe mogą się składać tylko z jednolitej, bezpostaciowej masy podstawowej (matriks), tworzonej przez albuminy, lub też zanurzone w niej mogą być globoidy i krystaloidy białkowe, mające zdolność pęcznienia (rącznik pospolity). Globoidy tworzy bezpostaciowa fityna, która składa się z nierozpuszczalnych soli kwasu fitynowego lub kwasu fitowego (inozytylosześciofosforowego). Ziarna aleuronowe mogą zawierać także enzymy hydrolityczne, np. proteazy i kwaśną fosfatazę (np. w nasionach grochu), niezbędne do gromadzenia białek i fityny oraz amylazę i rybonukleazy. Dzięki obecności tych enzymów w ziarnach aleuronowych odbywa się nie tylko gromadzenie i udostępnianie materiałów odżwyczych, ale także trawienie innych składników komórki. Ponadto uruchomienie alfa-amylazy w warstwie aleuronowej ziarniaków rozpoczyna proces kiełkowania. WYSTĘPOWANIE: Komórki wypełnione ziarnami aleuronowymi tworzą zewnętrzną warstwę bielma (tzw. bielmo aleuronowe) w ziarniakach traw zwaną warstwą aleuronową. Przy wyrobie białek mąki z ziarniaków zbóż warstwa aleuronowa wraz z okrywą owocowo-nasienną jest odrzucana jako tzw. otręby. Ziarna aleuronowe występują ponadto w zewnętrznej warstwie bielma w rodzinie rdestowatych oraz we wszystkich komórkach bielma u rącznika pospolitego, a także w liścieniach, np. grochu i fasoli.
65) Znormalizowany różnicowy wskaźnik wegetacji, NDVI (ang. Normalized Difference Vegetation Index) - jest to wskaźnik stosowany w pomiarach teledetekcyjnych (badania wykonywane z pewnej odległośc, izdalnie, przy wykorzystaniu odpowiednich czujników, wykonuje się je np. z samolotów, z przestrzeni kosmicznej lub z powierzchni ziemi) pozwalający określić stan rozwojowy oraz kondycję roślinności. NDVI bazuje na kontraście między największym odbiciem w paśmie bliskiej podczerwieni a absorpcją w paśmie czerwonym. Wskaźnik został zastosowany po raz pierwszy przez J.W. Rouse'a w 1973 roku i obliczany jest według wzoru:

Żywe, zielone rośliny pochłaniają promieniowanie słoneczne fotosyntetycznie czynne w zakresie 0,4-0,7 μm. Promieniowanie o falach dłuższych (0,7-1,1 μm) absorbowane jest w niewielkim stopniu. Wskaźnik przyjmuje wartości od -1 do 1. Wyższe wartości wskaźnika odpowiadają wyższemu odbiciu w zakresie podczerwieni i mniejszemu w zakresie czerwieni. Wysoka wartość wskaźnika odpowiada terenom pokrytym bujną roślinnością o dobrej kondycji. Wskaźnik NDVI stosowany jest do zbierania informacji o natężeniu fotosyntezy oraz do prognozowania plonów lub ilości biomasy wytworzonej przez ekosystem.
Brak komentarzy:
Prześlij komentarz