
Szparki mają istotne znaczenie dla funkcjonowania układu wentylacyjnego roślin pozwalającego na dostarczanie niezbędnego dla procesu fotosyntezy dwutlenku węgla. Dzięki nim rośliny mogą skutecznie pobierać ten gaz chroniąc się równocześnie przed szkodliwymi stratami wody w wyniku transpiracji. Funkcjonowanie szparek wiąże się ze zdolnością do zmiany kształtu komórek szparkowych. Komórki te poprzez zmianę turgoru, wykorzystując zjawisko osmozy, powodują zamykanie (obniżenie turgoru) lub otwieranie (podwyższenie turgoru) szparek. Charakterystyczną cechą budowy komórek szparkowych jest nierównomierna grubość i specyficzna struktura ich ścian, związana z układem mikrofibryli celulozowych. W zwykłych komórkach cylindrycznych mikrofibryle ułożone są poprzecznie w stosunku do głównej osi komórki wzdłuż jej podłużnej osi. W nerkowatych komórkach szparkowych mikrofibryle celulozowe rozchodzą się promieniście od otworu szparkowego. W związku z tym obwód szparki zostaje wzmocniony, a sama komórka wygina się na zewnątrz w trakcie rozwierania. U traw środkowe części komórek szparkowych oddalają się wówczas, gdy wzrasta objętość ich bulwkowatych zakończeń, zdolnych do rozciągania się, co pozwala na rozszerzenie szczeliny między komórkami. Dawniej sądzono, że zmiany turgoru komórek szparkowych są skutkiem hydrolizy i syntezy skrobi w chloroplastach tych komórek.
Obecnie wiadomo, że ruchy szparek powodowane są zmianą ciśnienia osmotycznego kationów potasu pobieranych przez komórki szparkowe przy jednoczesnym wydzielaniu protonów.
 |
| Źródło: organellum.blogspot.com - Proces zamykania i otwierania aparatu szparkowego. |
Pobieranie jonów K+ zależne jest od stężenia dwutlenku węgla wewnątrz liścia (od intensywności fotosyntezy, czyli pośrednio od światła, zaopatrzenia w wodę, temperatury). Transport jonów potasu do wakuoli zachodzi w wyniku aktywacji pompy elektrogenicznej. Światło niebieskie, absorbowane przez karotenoid zeaksantynę, znajdującą się w tylakoidach gran chloroplastu, aktywuje H+ - ATPazę w plazmolemie komórek szparkowych. Na skutek wzbudzenia zeaksantyny przez fotony światła niebieskiego zostaje zapoczątkowana kaskada zdarzeń, w wyniku których sygnał zostaje przekazany do cytoplazmy, gdzie aktywuje kinazę serynowo-treoninową. Kinaza fosforyluje C-końcowy fragment H+ - ATPazy, aktywując enzym. Przyłączenie małego (32 kDa) białka regulatorowego, 14-3-3, do ufosforylowanego końca C H+-ATPazy umożliwia stabilizację enzymu. Defosforylacja enzymu prowadzi do dysocjacji 14-3-3. H+ - ATPaza, wykorzystuje ATP wytwarza gradient protonowy przez błonę, co umożliwia transport K+ przez specyficzne kanały potasowe i akumulację tego kationu w wakuoli. Do wakuoli pobierane są też jony chloru (na drodze symportu Cl-/H+), co pozwala zobojętnić ładunek kationu nagromadzonego w wakuoli.
Innym czynnikiem wpływającym na otwieranie aparatów szparkowych są jony jabłczanowe. Anion jabłczanowy powstaje przez hydrolizę skrobi, natomiast Cl- jest transportowany z komórek sąsiednich. Wzrastające stężenie jonów obniża potencjał wodny komórki szparkowej wywołując napływ wody z komórek o większych potencjale, napływająca woda zwiększa turgor, duży turgor jest przyczyną wyginania się komórek szparkowych i otwierania szparek.
 |
Źródło: images.slideplayer.pl - "Fizjologia roślin. WODA - właściwości; Transport przez błony" - prezentacja użytkownika Elżbieta Ziomek - Rozkład skrobi i akumulacja jonów potasowych w cytoplazmie komórki szparkowej powoduje obniżenie potencjału wodnego tejże komórki, co powoduje napływ do niej wody z komórek o większym potencjale wodnym, które ją otaczają, co powoduje zwiększenie turgoru w komórce szparkowej, a to stanowi przyczynę wyginania się komórek szparkowych, co za tym idzie otwierania szparek.
|
Zamykanie się aparatów szparkowych następuje też pod wpływem innych związków chemicznych, m.in kwasu abscysynowego, którego stężenie w soku roślin rośnie znacznie w warunkach suszy.
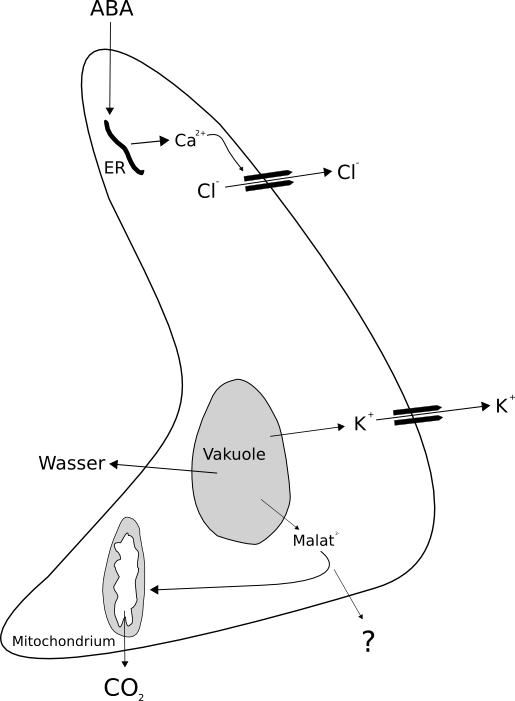 |
| Źródło: Wikipedia. |
Powyższy schemat ilustruje mechanizm zamykania aparatów szparkowych pod wpływem ABA. Kwas abscysynowy powoduje otwarcie kanałów jonowych i w następstwie utratę wody co prowadzi do zamknięcia aparatu szparkowego.
Brak komentarzy:
Prześlij komentarz