

EWOLUCJA I PRZEGLĄD SYSTEMATYCZNY KORZENI:
Rośliny naczyniowe istnieją co najmniej od syluru (okresu, który charakteryzował wzmożony wulkanizm, brak gleby oraz silne wiatry, które sprawiały, że lądy nie stanowiły zbyt gościnnego środowiska. Rośliny naczyniowe, które znane są od późnego ordowiku zaczęły z wolna zasiedlać bardziej przyjazne środowiska. Cieniutkie chwytniki zakotwiczały pierwsze z nich w cienkiej warstwie gleby. Później pojawiły się również korzenie i cały zestaw urządzeń niezbędnych do przetrwania na lądzie takich jak drewno, łyko oraz liście. Część roślin zaczęła wytwarzać osłonięte zarodniki zdolne przenosić się na duże odległości. Rośliny naczyniowe zaczęły oddalać się od swego pierwotnego środowiska - przybrzeżnych bagien i posuwać w głąb lądu). Z tego okresu pochodzą najstarsze zachowane skamieniałości takich organizmów. Nie jest jednakże pewne, czy te rośliny miały korzenie. Skamieniałości pozwalające potwierdzić istnienie struktur podobnych do korzeni pochodzą z dewonu. Prawdopodobnie korzenie wyewoluowały odrębnie u Lycopodiophyta (Widłaków) i Euphyllophyta (jest to klad obejmujący wszystkie rośliny nasienne i niemalże wszystkie rośliny zarodnikowe określane dawniej mianem paprotników. Grupa ta jest siostrzaną dla oddzielonej wcześniej linii rozwojowej widłaków. Obejmuje ona pochodzące od wspólnego przodka rośliny, których pozycję filogenetyczną i podział na główne linie rozwojowe przedstawawia powyższy kladogram (źródło: Wikipedia). Najstarsze znane organizmy z tej grupy to przedstawiciele kopalnego rodzaju Psilophyton występujący w dewonie. Nazwa grupy sugeruje, że do synapomorfii należy wykształcenie liści (makrofili) w przeciwieństwie do występujących u wcześniejszych linii rozwojowych listków (mikrofili będących wytworami epidermy). Z danych kopalnych wynika jednakże, że makrofile powstały niezależnie u dwóch głównych linii rozwojowych tych roślin, tj. odrębnie u paprotników z grupy monilofitów i odrębnie u roślin nasiennych. Za prawdziwe synapomorfie uznaje się wykształcenie korzeni o monopodialnym wzroście i endogenicznie powstających odgałęzieniach bocznych, wykształcanie zarodni wzdłuż krańcowych odcinków odgałęzień pędów i rozchodzenie się wzdluż tych krawędzi (cecha występująca u pierwotnych przedstawicieli, która później uległa rozmaitym modyfikacjom).
Rośliny naczyniowe istnieją co najmniej od syluru (okresu, który charakteryzował wzmożony wulkanizm, brak gleby oraz silne wiatry, które sprawiały, że lądy nie stanowiły zbyt gościnnego środowiska. Rośliny naczyniowe, które znane są od późnego ordowiku zaczęły z wolna zasiedlać bardziej przyjazne środowiska. Cieniutkie chwytniki zakotwiczały pierwsze z nich w cienkiej warstwie gleby. Później pojawiły się również korzenie i cały zestaw urządzeń niezbędnych do przetrwania na lądzie takich jak drewno, łyko oraz liście. Część roślin zaczęła wytwarzać osłonięte zarodniki zdolne przenosić się na duże odległości. Rośliny naczyniowe zaczęły oddalać się od swego pierwotnego środowiska - przybrzeżnych bagien i posuwać w głąb lądu). Z tego okresu pochodzą najstarsze zachowane skamieniałości takich organizmów. Nie jest jednakże pewne, czy te rośliny miały korzenie. Skamieniałości pozwalające potwierdzić istnienie struktur podobnych do korzeni pochodzą z dewonu. Prawdopodobnie korzenie wyewoluowały odrębnie u Lycopodiophyta (Widłaków) i Euphyllophyta (jest to klad obejmujący wszystkie rośliny nasienne i niemalże wszystkie rośliny zarodnikowe określane dawniej mianem paprotników. Grupa ta jest siostrzaną dla oddzielonej wcześniej linii rozwojowej widłaków. Obejmuje ona pochodzące od wspólnego przodka rośliny, których pozycję filogenetyczną i podział na główne linie rozwojowe przedstawawia powyższy kladogram (źródło: Wikipedia). Najstarsze znane organizmy z tej grupy to przedstawiciele kopalnego rodzaju Psilophyton występujący w dewonie. Nazwa grupy sugeruje, że do synapomorfii należy wykształcenie liści (makrofili) w przeciwieństwie do występujących u wcześniejszych linii rozwojowych listków (mikrofili będących wytworami epidermy). Z danych kopalnych wynika jednakże, że makrofile powstały niezależnie u dwóch głównych linii rozwojowych tych roślin, tj. odrębnie u paprotników z grupy monilofitów i odrębnie u roślin nasiennych. Za prawdziwe synapomorfie uznaje się wykształcenie korzeni o monopodialnym wzroście i endogenicznie powstających odgałęzieniach bocznych, wykształcanie zarodni wzdłuż krańcowych odcinków odgałęzień pędów i rozchodzenie się wzdluż tych krawędzi (cecha występująca u pierwotnych przedstawicieli, która później uległa rozmaitym modyfikacjom).
 |
| Źródło: Wikipedia. |
Poniższe ilustracje pochodzą z prezentacji: "Rhyniophyta"Štěpánka Bílková


Zróżnicowanie pomiędzy należącymi do Euphyllophyta roślinami zarodnikowymi a nasiennymi na etapie korzenia zarodkowego wskazuje na niezależną ewolucję korzeni u obu tych grup roślin. Rośliny zarodnikowe wykazują homoryzję (ang. homorrhizy - zjawisko wyrastania wszystkich korzeni z tego samego źródła, które charakterystyczne jest np. dla paprotników - nie wykształca się u nich korzeń zarodkowy, a wszystkie korzenie wyrastają z pędu u nasady pierwszego liścia. Homoryzja jest obserwowana także w rozwoju zarodka traw. Zarodek owych roślin wykształca wiele zaczątków korzeni. Pojęcie to zostało wprowadzone przez K. Goebela), rośliny nasienne zaś - alloryzję (ang. allorrhizy to zjawisko wykształcania korzeni niezależnie od pędu. Jest ono typowe dla roślin okrytonasiennych, u których w zarodku można wyróżnić wyraźny biegun korzeniowy. Jednakże u części jednoliściennych pomimo wykształcania zaczątków korzeni występuje tendencja do homoryzji).
POCHODZENIE KORZENI:
Korzenie są organem homologicznym do pędu. We wczesnym dewonie większość telomów była skierowana ku górze i pełniły funkcję organów przeprowadzających fotosyntezę, zaś pozostałe ustawione były poziomo, spoczywając na podłożu. Korzenie prawdopodobnie powstały z telomów grupy rugiej, które przystosowały się do pobierania wody oraz soli mineralnych, zapewniając jednocześnie roślinie zakotwiczenie. Na powstanie korzeni w wyniku rozdzielenia funkcji telmów wskazują zarówno analizy molekularne, genetyczne, jak i rozwojowe.
Pomimo niezależnej ewolucji korzenie niemalże wszystkich roślin wykształcają włośniki. Istnieje korelacja pomiędzy długością włośników a zdolnością do pobierania fosforanów. Około 80% roślin lądowych żyje w symbiozie z grzybami, tworząc mikoryzę. U tych gatunków włośniki są krótsze. Istnieją dowody wskazujące na istnienie mikoryzy już w dewonie. Brak zdolności do tworzenia mikoryzy jest zatem cechą wtórną, która pojawiła się u roślin, które rozwinęły długie włośniki. Mikoryza arbuskularna istniała jeszcze zanim rośliny lądowe wykształciły korzenie Cały rozwój ewolucyjny korzeni jest wynikiem koewolucji z grzybami mikoryzowymi. Ektomikoryza, mikoryza erikoidalna, mikoryza storczykowata oraz rośliny nietworzące mikoryzy pojawiły się w wyniku ewolucji w kredzie, gdy okrytozalążkowe uległy znacznemu zróżnicowaniu. Wcześniej korzenie zapewniały głównie współpracę z grzybami. Zwiększenie łącznej długości systemu korzeniowego, wykształcenie licznych i długich włośników oraz powstanie zaawansowanych systemów obrony chemicznej pozwoliły części gatunków na dalszy rozwój korzeni bez mikoryzy.
Geny umożliwiające tworzenie mikoryzy zostały wykorzystane do stworzenia symbiozy z bakteriami Rhizobium. Bakterie te odpowiedzialne są za powstawanie na korzeniach roślin motylkowatych brodawek korzeniowych i możliwość korzystania przez te rośliny z azotu atmosferycznego. Analiza genomu chloroplastowego wskazuje, że zdolność tworzenia brodawek przez korzenie przedstawicieli 10 rodzin okrytonasiennych pojawiała się w wyniku ewolucji tylko raz. Chociaż nie zachowały się skamieniałości korzeni z brodawkami korzeniowymi, prawdopodobnie ta forma symbiozy powstała wraz z grupą roślin ją tworzących we wczesnym trzeciorzędzie.
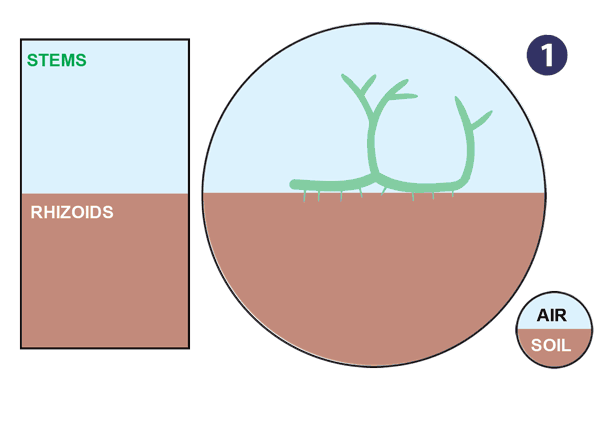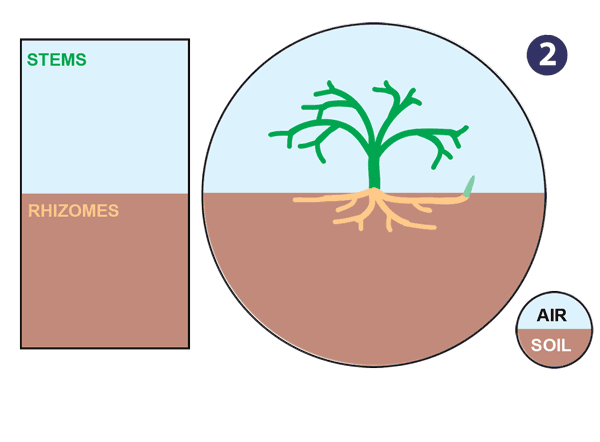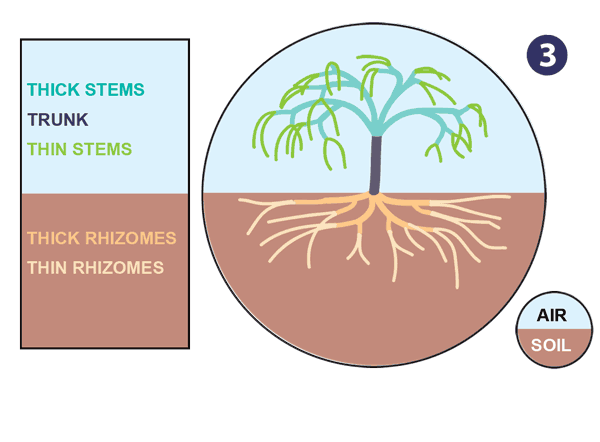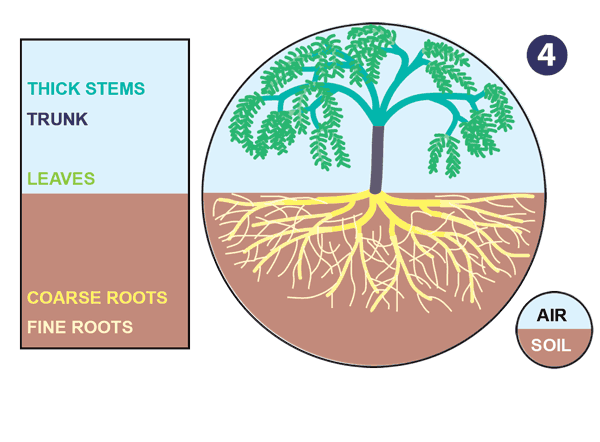 |
| Źródło: mycorrhizas.info. Ewolucja korzenia. |
- Aquatic plants without roots or leaves colonise land
- Early land plants with upright branches to intercept sunlight and horizontal stems (rhizomes) for propagation.
- Complex shoot and rhizome branching allows more effective use of resources and competition with other plants.
- Dimorphic subterranean rhizome systems evolve in response to allow more optimal nutrient and water uptake, mycorrhiza formation, and mechanical support. This occurs in parallel with the evolution of specialised aerial branches that are more efficient at photosynthesis.
- Some rhizomes become thinner and longer to increase contact with the soil and absorptive capacity, eventually becoming specialised as roots. These host mycorrhizal associations, grow faster and have a relatively short lifespan. In contrast, the thick rhizomes evolve protective features to limit permeability and facilitate long-term survival in soil.
Źródło: myccorhizas.info.
KORZENIE U POSZCZEGÓLNYCH GRUP SYSTEMATYCZNYCH:
- KORZENIE WIDŁAKOWCÓW (Lycopodiales) - wyrastają one z okolnicy (perycyklu), rzadziej ze śródskórni (endodermy). W obrębie pędu są one nierozgałęzione, ale po przebiciu się przez korę rozgałęziają się dychotomicznie (widlasto) lub także nieregularnie. Korzeń nie zawiera warstwy endodermy. W walcu osiowym najcieńszych korzeni ksylem ma budowę monoarchiczną (występuje jedno pasmo protoksylemu i protofloemu), zaś w grubszych diarchiczną (przy czym ksylem jest zwykle łukowato wygięty obejmując ramionami floem). Kora korzenia nie różni się od kory pędu. Na powierzchni występuje epiderma, z której komórek parami wyrastają włoski. Merystem wierzchołkowy opisywany był jako posiadający budowę warstwową, lecz wydaje się mieć jednak budowę nieuporządkowaną z wielką ilością komórek inicjalnych.
  |
| Źródło: http://phytoimages.siu.edu/ - Korzeń Lycopodium (Widłaka). |
 |
| Źródło: floweralley.files.wordpress.com. Korzenie Selaginella. |
- KORZENIE WIDLICZKOWCÓW (Selaginellales) - wyrastają one z charakterystycznych organów, które zwane są ryzomorfami (ryzomorfy to twory o sznurowatym lub korzeniastym pokroju wyróżniane u grzybów i widliczkowców. Nazwa ta pochodzi od dwóch łacińskich słów: rhiza - korzeń i morphe - kształt; u widliczkowców stanowią one specyficzne, bezlistne odgałęzienia pędów, wyrastające w dolnej ich części. Po zetknięciu z podłożem z owych odgałęzień wyrastają korzenie), gdy te dotkną podłoża. Jako jedyna grupa widłaków wytwarzają egzodermę z pasemkami Caspary'ego. Merystem wierzchołkowy ma strukturę nieuporządkowana, z niewyróżnialną komórką inicjalną lub z jedną taką komórką. Korzenie rozwidlają się dychotomicznie.

Źródło: http://www.biologydiscussion.com/

 |
| Źródło: http://www.biologydiscussion.com/ |
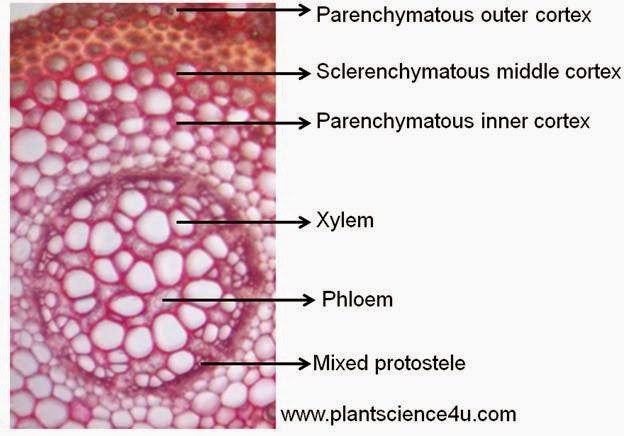
- KORZENIE PORYBLINOWCÓW (Isoetales) - są to korzenie wyrastające z rozwidlonych, dolnych płatów łodygi zwanych ryzoforami (dwóch lub czterech). Rozwijają się endogenicznie, przebijając grubą tkankę miękiszową i wyrastają w regularnych szeregach, równoległych do bruzd rozdzielających ryzofory. Co sezon wyrastają dwa, trzy takie szeregi, podczas gdy najstarsze korzenie znajdujące się na skrajach ryzofor odpadają wraz z fragmentem kory. Korzenie rozwidlają się dychotomicznie. Ich merystem wierzchołkowy ma budowę warstwową. W budowie wewnętrznej wyróżnia się położony ekstenctrycznie walec osiowy typu protosteli, otaczającą go endodermę i korę. W walcu osiowym występują pojedyncze pasma drewna i łyka, przy czym łyko położone jest po stronie kory pierwotnej (wiązki kolateralne). Kora wzdłuż łyka zwykle jest rozrywana, tak że w efekcie powstają na korzeniach szerokie przestwory powietrzne.

Źródło: herbaria.plants.ox.ac.uk - Korzenie Isoetales. - KORZENIE LEPIDOFITÓW (Lepidophytes) - korzenie przybyszowe tych roślin kopalnych wyrastały z rozległych kłączy wyrastających gwiaździście z podstawy pnia i rozgałęziających się dychotomicznie. Korzenie, zwane appendices, miały pochodzenie endogeniczne - ich kora stanowiła bowiem ciągłość z korą kłączy. Nie posiadały włośników i czapeczki. Osiągały długość do 40 cm i były nierozgałęzione, rzadziej dzieliły się dychotomicznie. Po obumarciu pozostawiały na kłączach charakterystyczne, ułożone skrętolegle, okrągłe i zgrubiałe blizny.

 |
| Źródło: Fossilhunters.xyz |
 |
| Źródło: http://peakgeology.blogspot.com/ |
*Lepidofity (łyszczydłowce/ lepidodendrowce) to rząd roślin kopalnych należących do klasy widłaków różnozarodnikowych. Przedstawiciele lepidofitów wymarły z końcem okresu karbońskiego. Lepidodendrony były widłakami dorastającymi do 30m wysokości i około 2m średnicy. Cechą, która łączyła je ze współczesnymi widłakami było to, że ich korona była rozgałęziona dychotomicznie. Były one okryte epidermą ze szparkami. Wyróżniał je ponadto charakterystyczne sposób przyrostu na grubość oraz wewnętrzna budowa - były one bowiem bylinami o zielnych i zielonych pniach. Pnie te miały organy korzeniotwórcze, zwane stigmariami. Sygilarie, podobnie jak lepidodendrony, osiągały znaczną wielkość. Miały proste, nierozgałęzione łodygi.
*Stigmaria - to dychotomicznie rozgałęziające się systemy korzeniowe kopalnych widłaków o pokrojach drzewiastych z rodzaju Lepidodendron i Sigillaria, tworzące szerokie podstawy utrzymujące w pionie rośliny na terenie bagiennym. Stigmarie najczęściej występują w postaci niewielkich spłaszczonych fragmentów i charakteryzują się skośnie położonymi owalnymi bliznami po korzeniach przybyszowych - apendyksach. Nagromadzenia stigmarii w położeniu przyżyciowym tworzą poziomy gleb kopalnych, zwanych ziemią (glebą) stigmariową, która ma lokalne znaczenie stratygraficzne. Odsłonięciem skał karbońskich ze stigmariami jest "skamieniały las" - Fossil Grove w Glasgow (Szkocja).
 |
| Źródło: hmag.gla.ac.uk |
Sygilarie posiadały organy korzeniotwórcze, również rozgałęziające się dychotomicznie, które zwane były Stigmariopsis i różniły się trochę wyglądem od stigmarii lepidodendronów. Od poziomo rosnących, płytko zanurzonych lub leżących na powierzchni Stigmariopsis odrastały prawdziwe korzenie o nazwie apendyks, po których zachowują się na stigmariach okrągłe, nieregularnie rozmieszczone blizny. KLIKNIJ I PRZECZYTAJ WIĘCEJ NA TEMAT SYGILARII.
Tak wyglądały lasy późnokarbońskie (dlaczego to NIE SĄ LASY możesz przeczytać tutaj, wraz z ciekawym opisem warunków klimatycznych panujących w owym ekosystemie: KLIK!)
Jeżeli chodzi o korzenie zwróć uwagę na to, że drzewa posiadają solidne korzenie, które głęboko zagłębiają się w ziemi (także wszerz) przez co nie są w stanie rosnąć na bagnach, bo duszone są przez wodę z braku powietrza. Byliny zaś mają kłącza, które rosną blisko lustra wody, gdzie powietrza jest w miarę dużo. Ich wątłe pędy, które bardzo szybko wyrosły są słabe i nawet niezbyt wielka wichura potrafiła je powalić co za tym idzie, że powstaje wolne miejsce i z kłączy rosnących w bagnistej ziemi wyrasta nowy pęd osiągając gigantyczne rozmiary. Zwróć zatem uwagę na to, że faktem jest, iż masa organiczna, która przyrasta w LESIE jest niewielka, szybko znika przez proces spróchniania. W karbonie natomiast z drzewiastych bylin (ale nie drzew!) ta masa przyrastała bardzo szybko; rośliny szybko rosły i równie szybko umierały - często pomagały im w tym wichury - kłącza zaczepione w niestabilnych bagniskach nie potrafiły utrzymać tak ogromnych, solidnych, majestatycznych pędów. Obalone pędy tonęły w bagnach, gdzie bez dostępu powietrza nie mogły się rozłożyć tylko pozostawały tam jako przyszły materiał na węgiel kamienny. Z kłączy zaś wyrastał nowy pęd, który szybko rosnąć dawał znowu dużą ilość niestabilnej masy zielonej i znów oczekiwał wichury, która spowoduje jego obalenie i wpadnięcie do czeluści bagna. Bagno pełniło tu rolę swego rodzaju konserwatora masy organicznej.
 |
| Źródło: images.zeno.org - Lepidodenrony, Sygillarie i Kalamity. Z lewej Lepidodenron. Zanurzone w wodzie są Kalamity. Sygillarie widoczne ze strony prawej. |
*Sygilarie (Sigillaria) - to wymarłe późnokarbońskie widłaki z rzędu lepidofitów. Sygilarie były widłakami dorastającymi do ponad 20 m wysokości i ok. 2 m średnicy u podstawy. Korona ich była nierozgałęziona lub rozgałęziona dychotomicznie na 2 lub 4 odgałęzienia. Liście były duże (do 1 m), płaskie, skupione w charakterystyczne dla sygilarii pióropusze. Pnie oraz gałęzie były okryte epidermą ze szparkami, nie zawierały wcale, lub bardzo mało, drewna. Wnętrze pni zaś często było puste. Wskazuje to, że sygilarie były bylinami o zielonych pniach.
- KORZENIE PSYLOTOWYCH (Psilotopsida) - przedstawiciele psylotowców (Psilotales) nie posiadają korzeni - podziemne kłącze pokryte jest chwytnikami, zaś w jego korze rozwijają się strzępki współżyjącego z rośliną grzyba. Podobnie współżyją z grzybami nasięźrzałowce (Ophioglossales), ale wytwarzają już korzenie, które zawierają jedną lub dwie wiązki otoczone grubą warstwą kory. Psylotowe stanowią najstarszą linię rozwojową w obrębie kladu monilofitów obejmującego współcześnie występujące rośliny zaliczane do skrzypów oraz paproci. Po uwzględnieniu badań molekularnych w filogenetyce tej grupy, zaliczono tu również nasięźrzałowce (Ophioglossales).
 |
| Źródło: http://www2.palomar.edu/ - Ophioglossales - widoczne korzenie. |
- KORZENIE SKRZYPOWYCH (Equisetopsida) - u wymarłych kalamitowatych (Calamitaceae) korzenie przybyszowe wyrastały licznie z węzłów podziemnych kłączy. W grubej korze zawierały liczne przestwory powietrzne, nie posiadały natomiast centralnego przewodu powietrznego znajdującego się w pniu nadziemnym. U skrzypów (Equisetum) korzenie także wyrastają z węzłów podziemnych kłączy, lecz są nieliczne, często pojedyncze i nierozgałęzione. Rzadko rozgałęziają się i na powierzchni okryte są brunatnymi włoskami tworzącymi pliśń podobną do tej pokrywającej podziemne kłącza.
*Kalamitowate (Calamitaceae) - to wymarła, głównie późnokarbońska rodzina skrzypowych. Najbardziej powszechnym przedstawicielem rodziny był rodzaj Calamites. Kalamitowate były roślinami o pokroju drzewiastym różnych rozmiarów. Największe z rodzaju Calamites dorastały do 30 m. Pień był wyraźnie segmentowany poprzecznie (tzw. węzły), podobnie do łodyg dzisiejszego skrzypu lub bambusa. Pień był również podłużnie żebrowany, a wewnątrz pusty. Miał grubą korę i grubą warstwę drewna wtórnego. Charakterystyczny dla wszystkich kalamitów jest przebieg wiązek przewodzących, które dochodząc do węzła rozwidlają się, co daje wrażenie jakby następny odcinek wiązki przewodzącej był przesunięty w węźle względem poprzedniej. Odróżnia to kalamitowate od archeokalamitów, które mają prostolinijny przebieg wiązki przez całą gałąź. Archeokalamity żyły w późnym dewonie i wczesnym karbonie - były podobne morfologicznie i anatomicznie do późniejszych kalamitów, lecz znacznie mniejsze od nich. Liście kalamitów były wąskie, wyłącznie w węzłach, zebrane w okółki. Kalamity posiadały podziemne kłącza, z których wyrastały korzenie.
WIĘCEJ INFORMACJI NA TEMAT KALAMITÓW ZNAJDZIESZ: TUTAJ.
 |
| Źródło: steurh.home.xs4all.nl - Korzenie Kalamitów. |
- KORZENIE STRZELICHOWYCH (Marattiopsida) - od innych paprotników wyróżniają się one dużą liczbą (ponad 12) skupień protoksylemu, ułożonych w walcu osiowym w formie aktynosteli. W korze korzeni znajdują się liczne przewody wydzielnicze, a także strzępki symbiotycznego grzyba - Strigeosporium marattiacearum (lęgniowce).
 |
| Źródło: plantsystematics.org - Przekrój poprzeczny korzenia charakterystyczny dla rodziny Marattiaceae. |
- KORZENIE PAPROCI (Polypodiopsida) - korzenie przybyszowe wyrastające z pędów, często u podstawy liści. Najsilniej rozwijają się u paproci drzewiastych, u gatunków zielnych często jest ich niewiele i są słabo rozgałęzione lub proste (do wyjątków należą tu długoszowate - Osmundaceae), a u paproci wodnych brak jest ich zupełnie (np. u salwinii pływającej funkcje korzenia przejmuje jeden z wyrastających w okółkach liści). U rozpłochowatych korzeni brak, zaś ich funkcję pełnią włośniki, które wyrastają wprost z kłącza.
 |
| Źródło: static.opracowania.pl - Widoczne są tu korzenie narecznicy samczej. |

Brak komentarzy:
Prześlij komentarz