Korzeń - organ dostarczający roślinom wodę i sole mineralne, cz. 2 - Systemy korzeniowe i modyfikacje korzeni.
Rozkład korzeni w podłożu uzależniony jest od wielu czynników oraz w sposób znaczący wpływa na poziom produktywności części nadziemnej rośliny.
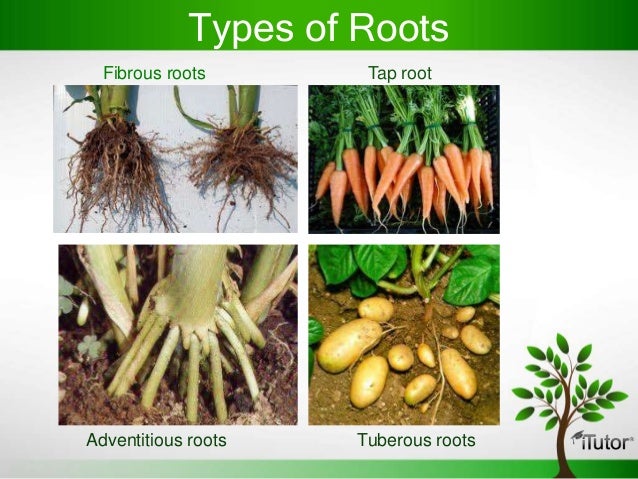
Z zawiązka korzenia wyrasta korzeń główny, który stanowi zazwyczaj przedłużenie pędu rosnącego pionowo w dół. Wzrost następuje w części wierzchołkowej, zaś boczne odgałęzienia, zwane korzeniami bocznymi, powstają wewnątrz korzenia głównego (endogenicznie). Korzenie boczne rosną ukośnie lub poziomo i rozgałęziają się dalej na korzenie boczne drugiego, trzeciego rzędu podobnie jak korzeń główny. System korzeniowy tak zbudowany nazywamy systemem osiowym, zaś jeżeli korzeń główny jest grubszy od innych, nazywany jest on systemem palowym. Gdy korzeń główny wcześnie przestaje rosnąć lub zanika - rozwija się pęk korzeni bocznych, ewentualnie pęk korzeni, które wyrastają wprost z łodygi (korzenie przybyszowe). System korzeniowy tego typu nazywamy wiązkowym. Ogólny schemat budowy systemu korzeniowego ulega różnym modyfikacjom funkcjonalnym u różnych grup ekologicznych roślin. W obrębie systemu korzeniowego zwykle istnieje zróżnicowanie anatomiczne, fizjologiczne oraz morfologiczne. Zróżnicowanie morfologiczne korzeni nosi nazwę heteroryzji [heteroryzja to zjawisko zróżnicowania korzeni na korzenie krótkie i długie. Korzenie krótkie zapewniają roślinom zaopatrzenie w wodę i sole mineralne. Zawierają one znaczne ilości tkanki miękiszowej, rosną wolno, zaś ich przyrost wtórny jest ograniczony. Korzenie długie rosną szybko i osiągają znaczną długość. Ich podstawową funkcją jest zapewnienie roślinie umocowania w podłożu a także transport substancji pobranych przez korzenie krótkie. U drzew, których system korzeniowy jest silnie rozbudowany korzenie długie dzielone są na korzenie pionierskie, tworzące główne gałęzie systemu oraz korzenie macierzyste, z których wyrastają korzenie żywiące (krótkie). Te ostatnie często mają mikoryzę].
KSZTAŁTY KORZENIA GŁÓWNEGO:
burakowaty - wyraźnie zgrubiały w pobliżu nasady, nagle zwężający się u dołu,
stożkowaty - najgrubszy u nasady, stożkowato zwężający się u dołu,
walcowaty - na całej długości ma zbliżoną średnicę,
wrzecionowaty - rozszerzony w części środkowej, zwężający się u dołu i góry.
Ze względu na sposób pobierania wody są one dzielone na systemy korzeniowe intensywne, ekstensywne oraz powierzchniowe:
System intensywny - występuje u jednoliściennych, zwłaszcza u traw i charakteryzuje się obecnością bardzo licznych oraz silnie rozgałęzionych korzeni przybyszowych. Korzenie takie pobierają duże ilości wody z niewielkiej objętości podłoża. Pomimo stosunkowo niewielkiej długości poszczególnych korzeni, ich sumaryczna długość może sięgać wielu kilometrów.
System ekstensywny - jest on typowy dla większości roślin dwuliściennych. Korzeń główny oraz korzenie boczne są tutaj miernie rozgałęzione, przerastają dużą objętość gleby, lecz wykorzystują ją w sposób mało ekonomiczny. Korzenie są długie i osiągają kilkadziesiąt metrów długości.
System powierzchniowy - jest on charakterystyczny dla sukulentów. Jest on rozległy, lecz bardzo płytki. Drobne korzenie w czasie suszy zasychają i wyrastają na nowo w przypadku zwilżenia podłoża.
Pomimo istnienia warunkowanego genetycznie generalnego układu korzeni, system korzeniowy wykazuje dużą plastyczność, dostosowując się do niejednorodnych warunków panujących w podłożu. W miejscach, gdzie ilość składników pokarmowych jest wyższa, następuje zagęszczenie systemu korzeniowego poprzez powstanie dodatkowych rozgałęzień bocznych. Rozwój systemu korzeniowego oznacza zwiększenie jego biomasy. Koszt rozbudowy systemu korzeniowego w kierunku zasobów substancji mineralnych musi być zrównoważony korzyściami z wchłonięcia dodatkowych soli mineralnych. W przeciwnym wypadku rozwój całego organizmu roślinnego zostanie zahamowany. Szereg doświadczeń na jęczmieniu zwyczajnym (Hordeum vulgare) wykazał, że przydział asymilatów niezbędnych do wydłużania i rozgałęziania korzeni zależy od dostępności fosforu oraz jonów NH4+ i NO3-, jednak nie zależy od zawartości jonów K+. Jednakże wyniki doświadczeń na innych gatunkach nie potwierdzają wniosków dla azotu. Prawdopodobnie gęstość systemu korzeniowego jest ważna dla jonów mało ruchliwych, takich jak fosforany i ma mniejsze znaczenia dla jonów łatwo przemieszczających się w glebie. Zrozumienie mechanizmów wpływających na rozwój systemu korzeniowego jest ważne ze względów naukowych, gospodarczych i środowiskowych.
Kluczowe dla przewidywania kształtu architektury systemu korzeniowego okazały się stężenia i transport wewnątrz organów hormonów roślinnych: auksyn, cytokinin, giberelin i kwasu abscysynowego. Najważniejsze w regulacji rozwoju korzenia okazały się auksyny. Druga grupa czynników to regulacja genetyczna. Duża część poznanych genów regulujących proces rozwoju systemu krzewionego to geny wpływające na syntezę hormonów roślinnych, bądź też kodujących receptory wrażliwe na owe hormony. Wzajemny wpływ na morfologię korzeni oraz grzybni istnieje u gatunków, które tworzą mikoryzę. Zjawisko to dotyczy 95% roślin. Między partnerem roślinnym oraz grzybowym istnieje dialog molekularny, który polega na wymianie informacji poprzez wydzielanie substancji sygnałowych. Rośliny informują o położeniu korzeni wydzielając związki należące do flawonoidów i polifenoli. Mniej jest wiadomo o sygnałach, które przekazywane są przez komponent grzybowy. Przypuszcza się, że grzyby stosują w komunikacji długodystansowej fitohormony, głównie cytokininy. Interakcje obejmują poza morfologią także wyłączenie reakcji odpornościowej korzeni roślin, syntezy fitoaleksyn [grupy organicznych związków chemicznych należących do izoflawonoidów, seskwiterpenów, poliacetylenów oraz innych grup wytwarzanych przez rośliny w odpowiedzi na atak patogenów. Zdrowe organy roślinne zawierają bardzo małe ilości fitoaleksyn. Ich stężenie wzrasta w wyniku działania czynników stresowych, takich jak zranienie, przechłodzenie, kontakt z patogenem lub jego produktami. Stężenie fitoaleksyn wzrasta w tkankach roślinnych wskutek synergistycznego działania kwasu jasmonowego (kwas jasmonowy to organiczny związek chemiczny będący przedstawicielem grupy regulatorów wzrostu i rozwoju roślin określanej jako jasmonidy. Struktura owych związków podobna jest do struktury prostaglandyn. Kwas jasmonowy to pochodna cyklopentanonu - powstaje w wyniku utleniania kwasu linolenowego. Kwas jasmonowy to inhibitor wzrostu oraz różnicowania komórek, a także stymulator końcowych etapów ontogenezy roślin czyli dojrzewania owoców czy też starzenia się liści. Kwas jasmonowy oraz jego pochodne, m.in. ester metylowy hamują wzrost pędów i korzeni roślin oraz fotosyntezę, modulują reakcje roślin na stres, wzmagają oddychanie komórkowe, a także mają wpływ na ekspresję genów) i etylenu] czy też chitynaz. Nie zostało rozstrzygnięte, czy grzyby mikoryzowe nie wywołują reakcji obronnej, czy też wyłączają ją, przesyłając odpowiedni sygnał.
Źródło: Atlasofscience.org
MONOCOT = JEDNOLIŚCIENNE EUDICOT = DWULIŚCIENNE WŁAŚCIWE ETIOLACJA. wypłonienie - procesy warunkujące powstawanie charakterystycznych cech u roślin rosnących w ciemności. U większości roślin okrytonasiennych w ciemności nie są syntetyzowane chlorofile, w etioplastach (etiolowanych plastydach) wykształcają się ciała prolamellarne, zamiast struktur lamellarnych. FLOOD/BURIAL - zalane wodą. Funkcje korzeni: Najważniejsze oraz najczęściej spotykane funkcje korzeni to dostarczanie roślinie wody wraz z substancjami odżywczymi czy też przytwierdzenie rośliny. W korzeniu zachodzi wiele reakcji ważnych dla metabolizmu rośliny. Szacuje się, że 15-27% substancji organicznych rośliny jest wytwarzanych w korzeniu. Zdolność wchłaniania wody mają głównie drobne młode korzenie. Korzenie z wtórnym przyrostem pełnią funkcję transportową, która jest szczególnie ważna w rozbudowanych systemach korzeniowych.
Do korzeni trafia od 5% do 20% węgla organicznego zasymilowanego w wyniku fotosyntezy. W korzeniu syntetyzowanych jest wiele substancji regulatorowych takich jak auksyny, cytokininy, gibereliny, kwas absycynowy, jasmoniany i poliaminy. W przypadku cytokinin i kwasu absycynowego korzeń jest głównym miejscem syntezy owych związków. Związki regulatorowe przenoszone są przez drewno do części nadziemnej wpływając na morfogenezę całej rośliny. Korzenie uczestniczą również w odbieraniu informacji ze środowiska, a także modyfikują środowisko glebowe poprzez dostarczanie do niego dodatkowych związków chemicznych. Statolity obecne w centralnych komórkach czapeczki korzeniowej umożliwiają odbiór informacji o kierunku działania siły ciążenia. Korzeń główny wykazuje geotropizm dodatni, korzenie boczne pierwszego rzędu diageotropizm albo plagiogeotropizm. Korzenie wyższych rzędów są ageotropiczne. Zwykle korzenie wykazują fototropizm ujemny oraz szereg chemotropizmów pozwalających na wzrost w kierunku zasobów niezbędnych dla roślin, najważniejszym z nich jest hydrotropizm. Eskudaty korzeniowe zmieniają rozpuszczalność soli mineralnych, wpływają pozytywnie lub negatywnie na rozwój innych organizmów obecnych w glebie, przede wszystkim bakterii, grzybów oraz owadów. Ważny ekologicznie jest również udział korzeni w tworzeniu związków mutualistycznych, głównie mikoryzy oraz brodawek korzeniowych.
U roślin wieloletnich korzenie pełnią również funkcję spichrzową, a u niektórych gatunków służą do rozmnażania wegetatywnego, rzadko odgrywają ważną rolę w układzie wentylacyjnym roślin lub nawet pełnią funkcję asymilacyjną.
Uczestniczenie korzeni w rozmnażaniu wegetatywnym jest stosunkowo rzadkie, w naturze ustępując wśród organów podziemnych rozłogom i kłączom. Rośliny potomne mogą powstawać z odrostów korzeniowych (odrosty lub odrośla to pędy rozwijające się z pąków śpiących lub przybyszowych na korzeniach lub w dolnej części pnia roślin drzewiastych), np. u osiki i robinii akacjowej; z bulw korzeniowych (np. dalia), czasem przystosowanych do rozmnażania w formie łatwo odpadających od rośliny macierzystej rozmnóżek (np. ziarnopłon wiosenny). Inaczej jest w przypadku działalności ogrodniczej, podczas której wykorzystywana jest powszechnie zdolność wytwarzania korzeni przybyszowych z fragmentów pędów do tworzenia sadzonek.
ZAOPATRYWANIE W WODĘ I SOLE MINERALNE:
Woda jest pobierana przez ryzodermę (skórkę korzenia), zwłaszcza włośniki. Następnie przez korę pierwotną przenoszona jest głównie drogą apoplastyczną. Przenikając przez cienkościenne komórki przepustowe w śródskórni i przez warstwę okolnicy trafia do naczyń i cewek, którymi jest transportowana w górę pędu. Ze względu na obecność w ścianach komórkowych śródskórni ligniny i suberyny przenoszenie wody odbywa się z udziałem symplastu. Transport wody w górę pędu następuje w wyniku parcia korzeniowego oraz transpiracji. Pierwszy z procesów umożliwia zaopatrzenie w wodę między innymi młodych, bezlistnych jeszcze roślin. W drugim wypadku woda parująca z powierzchni rośliny warunkuje istnienie ciągłego strumienia wody płynącego przez roślinę. Wraz z wodą roślina pobiera rozpuszczone w niej sole mineralne ważne dla jej metabolizmu.
Pobór wody powinien nadążać za jej utratą następującą głównie przez szparki i przetchlinki, bowiem rośliny zdolne są do wzrostu tylko przy dodatnim bilansie wodnym. W praktyce zarówno system pobierający wodę, jak i przewodzący są mało sprawne w stosunku do wymagań transpiracji, nawet gdy woda w siedlisku rośliny występuje obficie. U wielu roślin stwierdza się w ciągu godzin dziennych znaczny ubytek wody, sięgający 40% świeżej masy. Szczególnie niekorzystnym bilansem związanym z dysproporcją między rozwojem ulistnionego pędu i systemu korzeniowego cechują się rośliny młode. W miarę wzrostu systemu korzeniowego rośliny zwiększają odporność na susze. Ilość wody pobieranej i transpirowanej przez rośliny jest bardzo zmienna w zależności od ich wielkości i gatunku, np. kukurydza transpiruje 2 litry wody dziennie, zaś dąb już tylko 570 litrów.
Korzenie są pierwszym organem, do którego wnikają pobierane z wodą przez roślinę substancje. W niektórych przypadkach stwierdzono w nich akumulację metali ciężkich, co sugeruje istnienie mechanizmów, które zmniejszają ryzyko zatrucia rośliny. Również już w korzeniach następuje przekształcenie bardziej toksycznych form azotu (amonowych) w formy organiczne, podczas gdy mniej toksyczne, jak azotany są transportowane do kolejnych organów w niezmienionej postaci. Rośliny wodne pobierają swobodnie wodę z otoczenia. Wykształcenie korzeni umożliwiających pobieranie wody z gleby stanowi przystosowanie do warunków lądowych.
UMOCOWANIE ROŚLINY W PODŁOŻU:
Z nielicznymi wyjątkami (hydrofity - rośliny wodnopączkowe, których pączki odnawiające zimują w wodzie, które biernie pływają na powierzchni wody) rośliny są trwale przytwierdzone do gleby. Rozchodzące się we wszystkie kierunki, silnie rozgałęzione korzenie zabezpieczają roślinę przed wyrwaniem przez wiatr, przez ruchy cząstek podłoża (np. wydm), przed wypłukaniem przez wodę. System korzeniowy roślin drzewiastych rozwija się asymetrycznie, dostosowując się do przeważających dla lokalizacji kierunków wiatru. Duże znaczenia dla stabilności drzew ma głębokość, do jakiej docierają korzenie. Jednak ograniczeniem wzrostu w głąb podłoża jest często zwierciadło wód podziemnych. Korzenie kotwiczące drzewo w wyniku przyrostu wtórnego uzyskują zwiększoną gęstość w swej części górnej i w pionie. W efekcie układ zagęszczonych elementów korzenia przypomina na przekroju belkę T, po stronie zawietrznej. Po stronie nawietrznej dochodzi do zagęszczenia, które na przekroju przypomina belkę I. Rośliny drzewiaste o palowym systemie korzeniowym wzmacniają swoje korzenie głównie w wyniku drewnienia. Dodatkowo, aby zapewnić sobie mocowanie w podłożu mogą wytwarzać dodatkowe korzenie podporowe (np. kukurydza wytwarza krąg wzmocnionych korzeni przybyszowych). Dodatkowe korzenie tworzą naturalny system przyporowy.
GROMADZENIE SUBSTANCJI ZAPASOWYCH:
U roślin wieloletnich korzeń służy zwykle do magazynowania substancji pokarmowych przenoszonych z części pędowej w trakcie okresu wegetacyjnego, niezbędnych przy ponownym rozwoju pędu po okresie spoczynku. Funkcje spichrzowe pełni w korzeniu tkanka miękiszowa kory pierwotnej, drewna lub łyka. W skrajnych przypadkach wykształcają się korzenie specjalnie zmodyfikowane do pełnienia tej funkcji zwane spichrzowymi.
KORZENIE ODDECHOWE - jest to typ nadziemnych korzeni, doprowadzających powietrze do systemu korzeniowego rośliny. Są to wykazujące geotropizm ujemny i w efekcie wyrastające pionowo w górę odgałęzienia korzeni podziemnych rosnących równolegle do powierzchni gruntu. Korzenie takie występują zwłaszcza u roślin rosnących na podłożu grząskim i beztlenowym, zwłaszcza w strefie podzwrotnikowej i międzyzwrotnikowej, np. w namorzynach. Mogą osiągać wysokość nawet 1,5 metra. U różnych gatunków i w różnych warunkach rozwijają się w różnej liczbie: jedno drzewo z rodzaju Awicennia (rośliny te rosną na wybrzeżach wszystkich kontynentów w strefie międzyzwrotnikowej, na północy sięgając po południowe Chiny, Stany Zjednoczone oraz Egipt, a na południu po południową Brazylię, Australię i Nową Zelandię oraz południowe krańce Afryki) może wytworzyć około 10 tysięcy korzeni oddechowych. Cechują się one dobrze rozwiniętym miękiszem powietrznym (aerenchymą), którego cechą charakterystyczną jest silny rozwój przestworów komórkowych tworzących ciągły system kanałów powietrznych zapewniających przenoszenie tlenu, potrzebnego do procesów oddychania. Korzenie od zewnątrz posiadają liczne przetchlinki o prześwicie zbyt małym by przepuszczać wodę, ale wystarczającym do wymiany gazowej. Gdy korzenie są zalane podczas przypływu z przestworów międzykomórkowych pobierany jest tlen, wydzielany dwutlenek węgla łatwo rozpuszcza się w wodzie, tak że w efekcie wewnątrz korzenia powstaje podciśnienie. W efekcie podczas odpływu do wnętrza korzeni wessane zostaje powietrze. Niedotlenienie skutkuje upośledzeniem transportu węglowodanów z liści do tkanek niefotosyntetyzujących oraz zaburzeniem pobierania soli mineralnych. Pneumatofory mangrowców, wystające ponad powierzchnię gleby lub zastoiskowej wody, chronią tkanki korzenia przed niedotlenieniem i zapobiegają zachodzącym w takich sytuacjach procesom fermentacji.
Źródło: Wikipedia. Korzenie oddechowe awicenii w namorzynach.
Źródło: Wikipedia. Korzenie oddechowe cypryśnika błotnego (Aroretum w Kórniku)
Źródło: Wikipedia. Korzenie podporowe roślin namorzynowych.
KORZENIE PODPOROWE - stabilizują one roślinę rosnącą na grząskim podłożu. Są to korzenie przybyszowe, które ukośnie wrastają w ziemię, wyrastające z łodygi z różnych wysokości nad ziemią. Występują najczęściej u drzew i krzewów rosnących na terenach podmokłych, zwłaszcza w tropikalnych lasach deszczowych i namorzynach. W lasac tropikalnych umożliwiają stabilizację mechaniczną i szybki wzrost w warunkach zacienienia, gdy lokowanie zasobów w korzeniu podziemnym oraz pniu w początkowej fazie wzrostu jest utrudnione. U gatunków namorzynowych rozwój systemu korzeniowego w podłożu jest ograniczony ze względu na panujące w nim warunki beztlenowe. Częściowe odsłonięcie korzeni zapewnia lepszy dostęp do tlenu. Obecność korzeni podporowych pozwala osiągnąć relatywnie wysoki wzrost przy małej średnicy pnia. Zapewniają także kontynuowanie wzrostu na wysokość po częściowym zmieszczeniu drzewa. Kukurydza wykształca około 20-30 korzeni tworzących pierścień, wyrastających z podstawy łodygi i solidnie mocujących roślinę w podłożu. Korzenie te mogą znajdować się w całości w podłożu lub częściowo nad podłożem, jednak w obu przypadkach ich budowa jest identyczna. Szczególną formą korzeni podporowych są korzenie szkarpowe. Korzenie szkarpowe (bądź też korzenie skarpowe lub deskowe) to silnie spłaszczone korzenie rosnące poziomo od nasady pnia drzew. Ich początkowo koncentryczne przyrosty ulegają z czasem modyfikacji i z czasem rosną na grubość przede wszystkim ku górze tworząc w efekcie wąskie, prostopadłe do pnia i rozszerzające się ku dołowi listwy. Wykształcają się u drzew o płytkim systemie korzeniowym rosnących zwykle na siedliskach wilgotnych, słabo napowietrzonych lub ze składnikami pokarmowymi skupionymi w płytkiej warstwie powierzchniowej gleby. Korzenie te poprawiają statykę (równowagę układu pozostającego w spoczynku) drzew, zwłaszcza odporność na wpływ wiatru.
Źródło: Wikipedia. Korzenie szkarpowe.
KORZENIE SPICHRZOWE - pełnią one funkcję organów magazynujących substancje pokarmowe. Są one grube i mięsiste - charakteryzują się silnie rozrośniętym miękiszem spichrzowym w korzeniu głównym lub w korzeniach przybyszowych. Są często mylone z bulwami pędowymi (zwłaszcza powstającymi na podziemnych rozłogach jak u ziemniaka i słonecznika bulwiastego oraz z bulwami hipokotylowymi. W tym drugim przypadku funkcje spichrzowe pełni hipokotyl, który często łączy się z korzeniem spichrzowym (np. u marchwi zasadniczą część organu spichrzowego stanowi korzeń). W korzeniach spichrzowych funkcję magazynującą pełni część korzenia, która ulega zgrubieniu. U marchwi zgrubienie powstaje w wyniku rozbudowania warstwy łyka wtórnego. Warstwa ta składa się głównie z komórek miękiszowych, zawierających materiały zapasowe. Z kolei u rzodkiewki podczas przyrostu wtórnego korzenia rozbudowana zostaje warstwa drewna wtórnego. Także tutaj drewno składa się głównie z komórek miękiszowych z materiałami zapasowymi. W wyjątkowy sposób rozwija się spichrzowa część korzenia buraka. Przyrost wtórny rozpoczyna się od powstania warstwy kambium i wytworzenia pierścieni drewna i łyka. W trakcie przyrostu wtórnego powstają jednak kolejne warstwy kambium, na zewnątrz od pierwszego pierścienia. Warstwy te odkładają w obu kierunkach głównie komórki miękiszowe, jedynie w pewnych odstępach różnicują się wiązki łyka i drewna. Na przekroju zgrubiałego korzenia buraka widoczne są pierścienie wiązek przewodzących, powstałych z kilku warstw kambium. Część korzenia marchwi, rzodkiewki czy buraka położona poniżej zgrubienia, pełniącego funkcję organu spichrzowego, jest dość cienka i może sięgać nawet do 1 m głębokości. Właściwe bulwy korzeniowe obecne są np. u dalii, ziarnopłonu wiosennego, gółki i wielu innych storyczykowatych.
Źródło: flexiprep.com - Typy korzeni spichrzowych.
Źródło: slideshare. Plastydy i ich rola w korzeniach, w tym rola spichrzania substancji zapasowych: tłuszczy, białek, węglowodanów.
KORZENIE CZEPNE - korzenie czepne to korzenie przybyszowe, które wyrastają z łodygi pnączy (formy życiowej roślin o długiej, wiotkiej łodydze, wymagających podpór, by mogły się wspinać do góry, do światła; w strefie umiarkowanej występują one rzadko, lecz w tropikalnych lasach są bardzo czeste, w tym liczne o zdrewniałych łodygach, tzw. liany, które wykorzystując drzewa jako podpory oszczędzają konieczność wytwarzania silnych i grubych pni by wydostać się z ciemnego dna lasu tropikalnego ku słońcu) i ułatwiające wspinanie się po podporach. Korzenie takie obecne są na przykład u bluszczu pospolitego. Mają od 1 do 15 mm długości, są silnie rozgałęzione i wciskają się w szczeliny podpory. Włośniki wydzielają substancje o wyjątkowo dużej przyczepności. Odmienna jest także orientacja włókien celulozowych w ścianach komórkowych takich korzeni. Rozkład włókien zapewnia zwiększoną wytrzymałość mechaniczną. Jeżeli jednak wyrastający korzeń przybyszowy napotka podłoże nadające się do wzrostu, jego system staje się luźny jak u zwykłych korzeni. U figowca pnącego korzenie czepne inicjowane są parami, wyrastając po obu stronach wiązki przewodzącej. Podobnie jak u bluszczu liczne włośniki wytwarzają substancje białkowe i polisacharydowe, które po wyschnięciu zapewniają trwały przytwierdzenie do podpory.
Źródło: ogrodmarzen.net - Bluszcz pospolity (Hedera helix L.) dobrze porasta pnie drzew.
KORZENIE ROŚLIN PASOŻYTNICZYCH - korzenie roślin pasożytniczych (ssawki lub haustoria) wykształciły się u roślin pobierających substancje pokarmowe, wodę i sole mineralne z innych roślin (rośliny pasożytnicze, takie jak np. zaraz, kanianka, łuskiewnik) lub też pobierających jedynie wodę i sole mineralne a przeprowadzających fotosyntezę samodzielnie (półpasożyty, takie jak np. jemioła, pszeniec i świetlik). Półpasożyty korzeniowe wytwarzają słabo rozwinięty system korzeniowy z wyrostkami, które po zetknięciu z korzeniem odpowiedniego gospodarza wrastają do jego wnętrza. Tymczasem pasożyty korzeniowe mogą rozwinąć się tylko wtedy, gdy nasiono znajdzie się w bezpośrednim kontakcie z odpowiednim żywicielem. W przypadku pasożytów łodygowych, ich korzenie wrastają do warstwy korowej gospodarza i dlatego zwane są one korowymi. Inne pasożyty łodygowe tworzą na powierzchni pędu gospodarza korzenie zgrubiałe w formie przyssawki, zwane ekstramatrykalnymi, z których wyrastają ssawki wrastające do wnętrza żywiciela. Wzrost ssawek wewnątrz organizmu gospodarza jest możliwy dzięki wydzielaniu enzymów rozkładających blaszki środkowe i ściany komórkowe w jego tkankach. W zależności od gatunki rośliny pasożytniczej ssawki łączą się z ksylemem, floemem lub powierzchnią tkanki miękiszowej rośliny gospodarza. Możliwe są także kombinacje wymienionych połączeń. Bliski kontakt tkanek pasożyta i żywiciela umożliwia temu pierwszemu pobieranie roztworu przemieszczanego przez drzewo lub floem. Wykształcenie ssawek zapewnia fizjologiczny pomost między gospodarzem a żywicielem. Możliwe jest nawet powstanie międzygatunkowych plazmodesm. Obecność korzeni żywiciela wykrywana jest przez rośliny pasożytnicze w wyniku wydzielania do gleby strigolaktonów - substancji, które zwykle służą do komunikacji roślin z grzybami tworzącymi mikoryzę arbuskularną (tworzona jest ona przez grzyby rzędu Glomales z trawami, większością roślin uprawnych, drzewami, krzewami, takimi jak wiązy, klony, motylkowate. Struktury strzępkowe rozwijają się tu pomiędzy komórkami gospodarza, tworząc wypełnione materiałem zapasowym rozdęcia - tzw. wehikule. We wnętrzu komórek korowych korzeni, produkowane są silnie rozgałęzione struktury zwane arbuskulami).
Źródło: biologiaogul.blogspot.com - Nasada pędu jemioły (Wscum sp.) z korzeniami wrośniętymi w gałąź drzewa żywicielskiego.
KORZENIE ASYMILACYJNE - korzenie asymilacyjne to korzenie przybyszowe, które przejęły funkcję organów asymilacyjnych. Najczęściej są one spotykane u epifitów (form wykorzystujących inne rośliny jako podłoże), np. u przedstawicieli storczykowatych. Zdolność przeprowadzania fotosyntezy mogą wykazywać także pneumatofory mangrowców oraz korzenie podporowe palm. Zawartość chlorofilu w korzeniach asymilacyjnych 12 zbadanych gatunków storczykowatych wyniosła od 30% do 119% jego zawartości w liściach. Najczęściej jednak wydajność fotosyntezy netto dla korzeni nie osiąga wartości dodatnich. Korzenie storczykowatych zwykle przeprowadzają fotosyntezę typu CAM. Rośliny z rodzaju zasennikowatych oraz przedstawiciele storczykowatych z rodzajów Campylocentrum, Dendrophylax, Taenophyllum są bezlistni, a korzenie przejęły funkcję dostarczania tym roślinom związków organicznych. Korzenie asymilacyjne mogą być taśmowate lub płatowate.
Źródło: wikimedia.org - Korzenie powietrzne storczykowatych często pełnią funkcję organów asymilacyjnych.
KORZENIE KURCZLIWE - występują one u roślin kiełkujących na powierzchni ziemi z nasion, których pędy (kłącza, cebule, skrócone łodygi) rozwijają się pod powierzchnią gruntu. Umożliwiają one roślinom wytwarzającym takie organy umieszczenie ich na odpowiedniej głębokości w glebie, tak aby w stanie spoczynku [w fizjologii spoczynek to stan azahamowania wzrostu, silnego ograniczenia procesów metabolicznych przy jednoczesnym zachowaniu pełnej zdolności do życia; rośliny wykształcają specjalne struktury spoczynkowe, które umożliwiają im przetrwanie w niesprzyjających warunkach środowiska. Struktura spoczynkowa u roślin wieloletnich może przyjmować postać kłącz, bulw, cebul, pędów u drzew i krzewów zaś postać pąków śpiących. W stan spoczynku zapadać mogą również całe rośliny. U roślin jednorocznych funkcję utworu spoczynkowego pełnią nasiona. W strefie klimatu umiarkowanego wiele roślin wchodzi w stan spoczynku zimą; w celu zabezpieczenia przed zamarzaniem drzewa i krzewy zrzucają liście i ograniczają tempo metabolizmu. Kłącza, bulwy oraz cebule przechodzą w stan spoczynku jesienią w wyniku ochłodzenia oraz sk®ócenia długości dnia. Ustępowanie spoczynku owych struktur następuje zwykle po okresie temperatur około 5 stopni Celsjusza. Jednakże w wielu przypadkach wystarcza okres tylko niewielkiego spadku temperatury w zakresie 5-20 stopni Celsjusza] mogły spokojnie przetrwać zimę, okres suszy lub gorąca. Ruch organu przetrwalnikowego, w wyniku wciągania go przez korzenie kurczliwe, może być arejestrowany dzięki wykonywanym cyklicznie fotografiom. W jednym sezonie wegetacyjnym roślina może zagłębić się o 20 cm. Korzenie kurczliwe występują u okrytonasiennyhc oraz niektórych gatunków paprotników. Stanowią u takich roślin jedynie część ich ogólnej liczby korzeni. Skurcz jest efektem zmian w komórkach miękiszowych kory pierwotnej. Komórki te wydłużają się radialnie, jednocześnie zmniejszając swoją długość wzdłuż osi korzenia. Zewnętrzne warstwy kory pierwotnej oraz epidermy nie zmieniają kształtu i w efekcie zmian w głębszych warstwach powstają u nich fałdy. Zagęszczeniu ulegają także tkanki steli. Przyjmuje się, że reakcje w obrębie steli mają pasywny charakter. U niektórych roślin, np. u lilii wszystkie korzenie mają zdolność do kurczenia się w
pewnym zakresie, u innych np. krokus (rys. A) tylko niektóre korzenie są
wyspecjalizowane w tym kierunku.
KORZENIE POWIETRZNE - są to korzenie spotykane u roślin rosnących w klimacie tropikalnym, głównie u epifitów. Służą one do pobierania wody z opadów i pary wodnej. U figowców korzenie powietrzne po zetknięciu z gruntem rozpoczynają przyrost wtórny na grubość, pojawia się w nich drewno tensyjne i stają się korzeniami podporowymi. Korzenie powietrzne storczykowatych, arekowatych, liliowatych, pochrzynowatych, krąpielowatych, amarylkowatych i komelinowatych pokryte są welamenem, tworzonym przez jedną lub wiele warstw komórek. Tkanka ta składa się z martwych komórek, które mają przekrój poprzeczny wielokątny, eliptyczny lub prostokątny. Zewnętrzne komórki egzodermy nie mają protoplastów, zaś ściany komórkowe ulegają lignifikacji. Jednakże na ogół komórki kory pierwotnej są żywe i zawierają chloroplasty. Stela ma niewielką średnicę. Otaczająca wiązki przewodzące endoderma składa się ze ściśle przylegających do siebie komórek o grubych ścianach i komórek przepustowych o cienkich ścianach zlokalizowanych w pobliżu biegunów protoksylemu. Budowa welamenu może ułatwiać kondensację pary wodnej.
Źródło: tuznajdziesz.pl - Korzenie powietrzne storczyków z rodzaju Vanda.
*Welamen (łac. velamen = okrycie, suknia) - jest to gąbczasta warstwa obumarłych komórek skórki na korzeniach powietrznych epifitów z rodziny storczykowatych i obrazkowatych. Służy ona do gromadzenia i wychwytywania wody z atmosfery i przytwierdzania roślin do nierówności pni. Gruba warstwa pustych komórek ma barwę srebrzystą w stanie suchym. Korzenie z welamenem nasączone wodą stają się zielone. U niektórych gatunków warstwa komórek welamenu jest postrzępiona w formie filcowato zbitych włosków (np. storczykowate z rodzaju pafiopedilum).
Źródło: CarolinaBiologicalSupplyCompany. Korzeń powietrzny Storczyka - budowa anatomiczna.
KORZENIE KORALKOWE - korzenie koralkowe występują u sagowców (monotypowej klasy roślin należącej do gromady nagonasiennych obejmującej jeden rząd sagowce (Cycadales) z dwiema rodzinami, 11 rodzajami i około 320-340 gatunkami. Naturalny zasięg i rozpowszechnienie tychże roślin jest współcześnie ograniczone, jednakże przodkowie sagowców podobni do żyjących obecnie odgrywali ogromną rolę w krajobrazie i stanowili podstawowe źródło pokarmu dla wielu dinozaurów). Wyrastają nad powierzchnię zapewniając siedlisko sinicom, zdolnym do wiązania azotu atmosferycznego. Sinice Nostoc i Anabaena zasiedlają korzenie boczne wyrastające z perycyklu i niewykazujące geotropizmu.
Źródło: Wikipedia.
Źródło: Wikipedia.
W dojrzałym korzeniu krótkie hormogonia [ruchliwki - wielokomórkowe twory występujące u sinic, służące do rozmnażania wegetatywnego, które są zdolne do aktywnego ruchu. Znajdują się wewnątrz osłonek zwanych pochwami, z których wypełzają i przekształcają się w nowe fragmenty nici sinic. Mogą otoczyć się grubymi ścianami komórkowymi i przybrać przetrwalnikowy charakter] dostają się przez tkankę okrywającą do przestrzeni w korze wtórnej. Tam zostają pokryte warstwą śluzu. Mające zdolność przeprowadzania fotosyntezy w korzeniach sagowców sinice wykazują miksotrofizm. Obecność symbiontów umożliwia roślinom korzystanie z azotu atmosferycznego. W przypadku Macrozamia riedlei wyliczono, że dzięki sinicom do ekosystemu trafia rocznie dodatkowo 18,6-18,8 kg azotu/ha. Azot organiczny jest przekazywany przez sinice roślinie w postaci cytruliny, glutaminy i w mniejszej ilości kwasu glutaminowego.
KORZENIE PROTEIDOWE - korzenie proteidowe zostały opisane w roku 1960. Składają się one z korzenia głównego o wyjątkowo licznych, jednakże krótkich korzeniach bocznych Ich występowanie początkowo stwierdzono również u roślin z rodziny Proteaceae, stąd nazwa korzeni. Później korzenie o podobnej budowie zostały wykryte także u roślin z rodzin: Betulaceae, Casuarinaceae, Elaeagnaceae, Mimosaceae, Fabaceae, Moraceae i Myricaceae. Korzenie te są adaptacją do warunków niedoboru fosforu, żelaza oraz azotu. Liczne korzenie boczne mogą zwiększać powierzchnię chłonną w stosunku do typowego korzenia 140-300 razy. Poprawa wydajności pobierania związków mineralnych jest również związana z wydzielaniem przez korzenie karboksylanów, fenoli i wody. Wydzieliny zwiększają bowiem rozpuszczalność trudno przyswajalnych składników odżwyczych. Wyróżniane są dwa główne typy korzeni proteidowych. Typ pojedynczy, gdy korzeń główny i liczne korzenie boczne mają kształt szczotki do mycia butelek oraz typ złożony, gdy roślina wykształca wiele rozgałęzionych korzeni tworzących zwartą matę w organicznej warstwie gleby.
Źródło: researchgate.net - Korzenie proteidowe u gatunków z rodzaju Proteaceae. link do publikacj "Metabolic adaptations of the non-mycotrophic Proteaceae to soil with a low phosphorus availability" - Hans Lambers : klik.
WIĘCEJ INFORMACJI NA TEMAT KORZENI PROTEIDOWYCH - KLIK.
KORZENIE MARCHEWKOKSZTAŁTNE: korzenie marchewkokształtne (ang. dauciform) to oddzielna forma morfologiczna korzeni występujących wśród Cyperaceae. Ich nazwa pochodzi od łacińskiej nazwy marchwi - Daucus. Korzenie te wykształcają krótkie korzenie boczne z licznymi, długimi włośnikami. Do powstania korzeni tego typu dochodzi w warunkach niedoboru fosforu. Pod względem funkcjonalnym są analogiczne do korzeni proteoidowych, jednakże strukturalnie są inne. Poza szczególną morfologią korzenie marchewkokształtne charakteryzują się wydzielaniem do podłoża znacznych ilości kwasów karboksylowych i fosfataz, które zwiększają rozpuszczalność dostępnych w glebie związków fosforu.
























 Źródło: Wikipedia.
Źródło: Wikipedia.
Brak komentarzy:
Prześlij komentarz