
I. Tkanka mięśniowa (łac. textus muscularis) - to jedna z podstawowych tkanek zwierzęcych. Składa się ona z włókien mięśniowych, zbudowanych z miocytów (zespołów komórek mięśniowych), posiadających zdolność do aktywnego kurczenia się.
Rodzaje tkanki mięśniowej to:
- tkanka poprzecznie prążkowana szkieletowa,
- tkanka poprzecznie prążkowana serca,
- tkanka gładka
 |
| Źródło: YouTube, film: "Skeletal Muscle Tissue". |
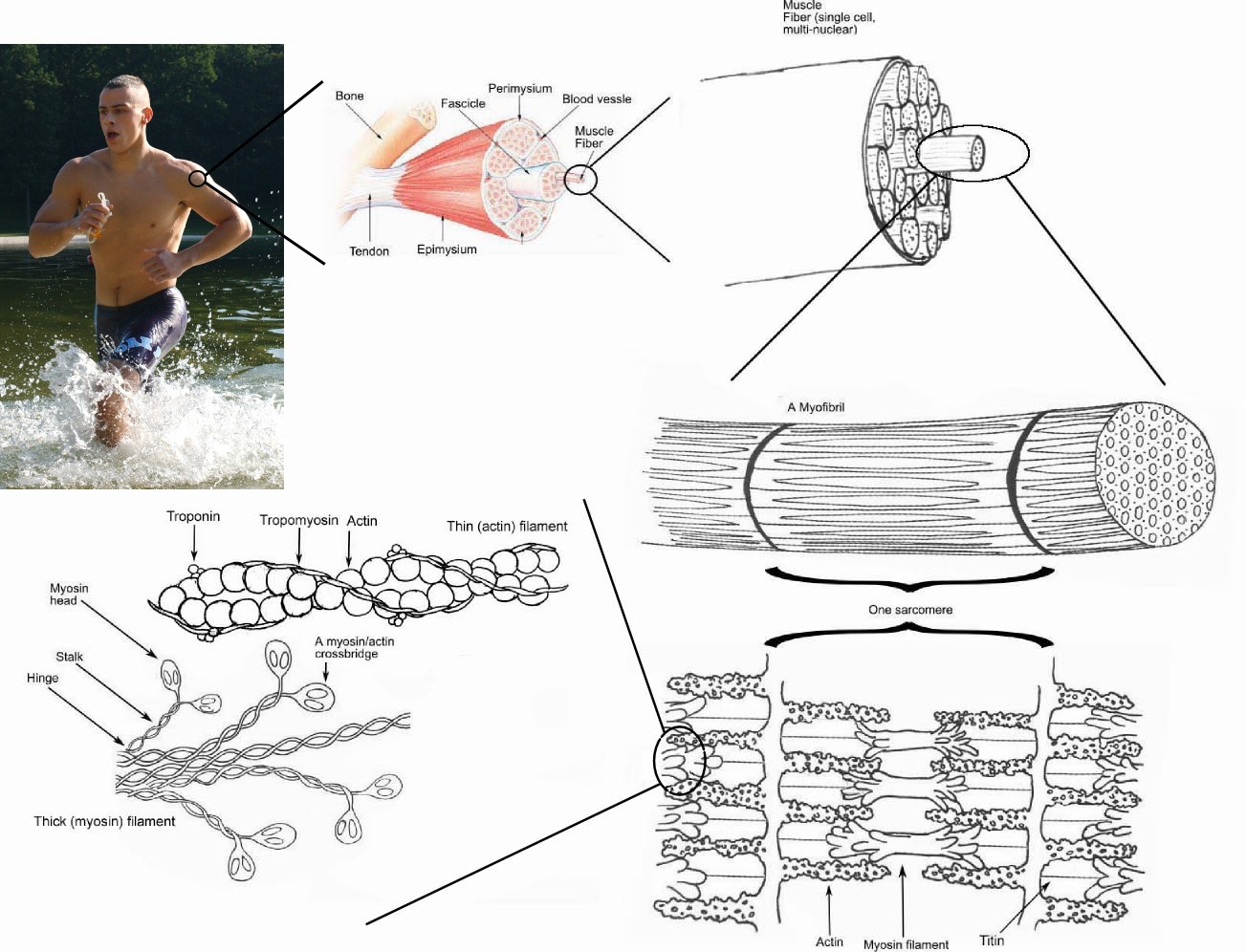 |
| Źródło: Wikipedia. Budowa mięśnia. |
Wykonanie skurczu następuje dzięki występowaniu w nich miofibryli, czyli włókienek kurczliwych zbudowanych z łańcuchów polipeptydowych. Efektywność ruchu w mięśniach jest możliwa dzięki ścisłemu ułożeniu włókien mięśniowych, pomiędzy którymi nie występuje żadne inna tkanka. Mechanizm działania miofibryli jest aktualnie przedmiotem dyskusji naukowej i istnieją na ten temat dwie rozbieżne teorie. Tkanka mięśniowa nie ma własnej substancji międzykomórkowej, a elementy mięśniowe połączone są ze sobą za pomocą tkanki łącznej wiotkiej. Pomimo obecności w komórkach mięśniowych jądra komórkowego oraz pewnego zdolności do podziału, ubytki w tkance mięśniowej tylko w niewielkim stopniu są uzupełniane w wyniku podziału nieuszkodzonych komórek. Najczęściej zostają one zastąpione tkanką łaczną tworzącą w tym miejscu bliznę. Tkanki mięśniowe, poprzecznie prążkowana serca i gładka unerwione są przez układ współczulny i działają niezależnie od woli człowieka. Natomiast mięśnie poprzecznie prążkowane, unerwione somatycznie, kurczą się zgodnie z wolą człowieka.
1) Tkanka poprzecznie prążkowana szkieletowa - elementami strukturalnymi, z których zbudowany jest ten typ tkanki, są komórki wielojądrzaste, nazwane włóknami mięśniowymi. Włókno mięśniowe ma więc charakter syncytium, które powstało w wyniku zespolenia wielu komórek. Dlatego też w każdy włóknie występuje od kilkudziesięciu do kilkunastu jąder, które położone są na obwodzie komórki, pod błoną sarkoplazmatyczną. Włókna mięśniowe mają kształt walcowaty, długość ich sięga od 1 do 5 cm, niekiedy zaś nawet do kilkunastu centrymetrów.
 |
| Źródło: teachcing.ncl.ac.uk - Mięśnie szkieletowe. |
Wnętrze włókna wypełniają prawie całkowicie włókienka kurczliwe (miofibryle). Biegną one równolegle do siebie, wzdłuż długiej osi włókna, najczęściej zebrane w pęczki, odizolowane skąpą ilością sarkoplazmy. Sarkoplazma, tj. cytoplazma komórek mięśniowych, zawiera czerwony barwnik - mioglobinę oraz znaczne ilości ziaren glikogenu. W komórkach tkanki mięśniowej znajdują się liczne mitochondria zwane również sarkosomami, słabo rozwinięty układ Golgiego, zlokalizowany w pobliżu jądra oraz siateczka śródplazmatyczna gładka. Siateczka śródplazmatyczna wystepuje w bezpośrednim sąsiedztwie włókien kurczliwych, tworząc bardzo regularny i skomplikowany układ kanalików podłużnych i poprzecznych. Kanaliki podłużne są elementami sieci sarkoplazmatycznej i noszą nazwę sarkotubul. Sarkotubule rozszerzają się na obu końcach sarkomeru tworząc cysterny, które sąsiadują z poprzecznie leżącymi kanalikami utworzonymi w wyniku wypuklenia się sarkolemmy - są to tzw. kanaliki pośrednie T. Do kanalików T przylegają cysterny sąsiadujących kanalików siateczki śródplazmatycznej tworząc tzw. triady. Za pośrednictwem tego systemu kanalików odbywa się wymiana substancji między miofibrylami a środowiskiem zewnętrrznym, przewodzenie bodźców skurczowych oraz transport jonów wapnia, niezbędnych do skurczu włókien mięśniowych.
Włókna mięśniowe dzieli się pod względem morfologicznym i czynnościowym na dwa podstawowe typy:
- włókna typu I - wolnokurczące się (zwane też z ang. slow twitching "ST"),
- włókna typu II - szybkokurczące się (fast twitching "FT")
Włokna wolnokurczące się zawierają wiele mitochondriów i duże stężenie mioglobiny (stąd zwane są też czerwonymi), co jest istotne, gdyż energię do skurczu czerpią z procesów tlenowych. Charakteryzują się one powolnym narastaniem siły skurczu i dużą wytrzymałością na zmęczenie.
 |
| Źródło: workout-polska.pl - Rodzaje włókien mięśniowych mięśni poprzecznie prążkowanych szkieletowych. |
Włókna szybkokurczące się (białe) zawierają mniejsze stężenie mioglobiny, kurczą się szybciej, ale są mniej wytrzymałe. Biorąc pod uwagę głównie źródła energii z jakich korzystają, wyróżnia się wśród nich:
- włókna typu IIA - glikolityczno-tlenowe, wykorzystujące energię wytworzoną w procesie glikolizy w cytoplazmie oraz w procesie fosforylacji oksydacyjnej w mitochondriach,
- włókna typu IIB - glikolityczne - korzystające głównie z energii wytworzonej podczas glikolizy - liczba mitochondriów jest w nich mniejsza.
Mięśnie człowieka zawierają oba rodzaje włókien, a ich wzajemny stosunek jest różny u różnych ludzi. U sportowców uprawiających dyscypliny siłowe przeważają włókna typu białego. Trening wytrzymałościowy powoduje zwiększenie potencjału tlenowego mięśni przez zwiększenie liczby naczyń kapilarnych w mięśniach.
Budowa włókienek kurczliwych - miofibryli jest bardzo złożona. Nie posiadają one jednorodnej struktury, lecz składają się z jaśniejszych oraz ciemniejszych odcinków, leżących na przemian. jaśniejsze odcinki są zbudowane z substancji, która pojedynczo załamuje światło - są to tzw. prążki izotropowe I, prążki ciemniejsze izotropowe jak i anizotropowe leżą we wszystkich miofibrylach na długiej osi włókna mięśniowego, wskutek czego powstaje wrażenie poprzecznego prążkowania całego włókna.
 |
| Źródło: atlas.histologiczny.cm.umk.pl - Tkanka poprzecznie prążkowana szkieletowa. Przekrój poprzeczny. |
 |
| Źródło: medcell.med.yale.edu - Tkanka mięśniowa gładka. |
 |
| Źródło: static.scholaris.pl - Komórki tkanki mięśniowej gładkiej o kształcie wrzecionowatym. |
 |
| Źródło: previews.123rf.com - Skurcz mięśni gładkich. |
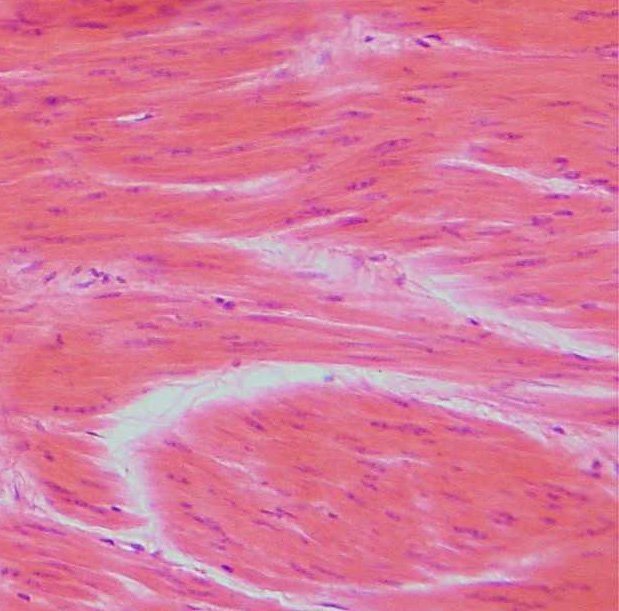 |
| Źródło: Wikipedia. Mięsień gładki pod mikroskopem. |
3) Tkanka mięśniowa poprzecznie prążkowana typu sercowego - jest to rodzaj tkanki mięśniowej występujący jedynie w sercu kręgowców. Mięsień sercowy różni się od mięśnia poprzecznie prążkowanego typu szkieletowego tym, iż włókna w mięśniu sercowym są widlasto rozgałęzione zaś jądra komórkowe ułożone są centralnie, podczas gdy w mięśniu poprzecznie prążkowanym włókna są złączone, a jądra znajdują się na obrzeżach włókien. Komórki tkanki sercowej człowieka są jedno, rzadko dwujądrzystymi komórkami, które wykazują poprzeczne prążkowanie.
 |
| Źródło: atlas.histologiczny.cm.umk.pl |
Mówi się, że jest to jedyny mięsień poprzecznie prążkowany, którego praca nie podlega woli (jednak pośrednio możemy wpływać na skurcze). Jest jednakże również kilka innych mięśni, które również są niezależne od woli: mięśnie ucha środkowego; mięsień kulszowo-jamisty; mięsień opuszkowo-gąbczasty; niektóre komórki mięśniowe mięśni oddechowych.
 |
| Źródło:YouTube, film: "Cardiac Muscle Tissue". |
II. Tkanka nerwowa (textus nervosus) - jest to utworzona przez neurony (komórki nerwowe) i komórki glejowe tkanka, która tworzy układ nerwowy. Odbiera, przekazuje i reaguje na bodźce pochodzące ze środowiska zewnętrznego, jak na przykład dotyk, temperatura czy światło. Przewodzi impulsy z neuronu do efektorów, od receptorów, przetwarza impulsy w adekwatne odpowiedzi,przewodzi impulsy z neuronu do innego neuronu, wytwarza substancje przekaźnikowe. Komórki nerwowe umożliwiają organizmowi normalne funkcjonowanie w danym środowisku, adekwatną odpowiedź w zależności od sytuacji w środowisku zarówno wewnętrznym organizmu jak i zewnętrznym stanie otoczenie, przez co przygotowują organizm do adekwatnej reakcji. Do neuronów należy również koordynacja aktywności intelektualnej, świadomości, podświadomości, aktywności ruchowej czy też czynności gruczołów dokrewnych. Tkanka nerwowa ma bardzo słabe możliwości regeneracyjne, jest także szczególnie wrażliwa na brak tlenu.
Tkanka ta złożona jest z licznych komórek nerwowych - neuronów. Składają się one z ciała komórki zawierającego jądro. Od tego ciała odchodzą krótkie wypustki zwane dendrytami oraz najczęściej jedna, długa i rozgałęziona na końcu wypustka - neuryt. Mogą go otaczać osłonki mielinowe. Dendryty odbierają bodźce i przekazują je do ciała komórki nerwowej, a stąd przez neuryt informacja trafia do następnej komórki nerwowej. Dzięki dendrytom i neurytom komórki nerwowe mogą spełniać swoje funkcje, czyli odbierać i przekazywać bodźce ze środowiska zewnętrznego i wewnętrznego do centralnego układu nerwowego - mózgu i rdzenia kręgowego. Centralny układ nerwowy, od którego odchodzą liczne włókna nerwowe pełni nadrzędną funkcję w stosunku do innych układów i całego organizmu. Układ nerwowy scala, kontroluje wszystkie czynności życiowe i funkcjonowanie żywego organizmu. Z tkanki nerwowego zbudowane są: ośrodkowy układ nerwowy: mózg i rdzeń kręgowy i obwodowy układ nerwowy.
KOMÓRKI NERWOWE:
NEURON (Komórka nerwowa) - to rodzaj elektrycznie pobudliwej komórki zdolnej do przetwarzania i przewodzenia informacji w postaci sygnału elektrycznego. Neurony są podstawowym elementem układu nerwowego zwierząt. Najwięcej neuronów znajduje się w ośrodkowym układzie nerwowym w skład którego wchodzi mózgowie oraz rdzeń kręgowy.
Typowy neuron jest zbudowany z ciała komórki (perikarion) oraz odchodzących od niego wypustek: aksonu i dendrytów. Neurony kontaktują się między sobą poprzez synapsy tworząc sieci neuronowe. Informacje od innych neuronów są odbierane przez synapsy położone na dendrytach, przewodzone wzdłuż neuronu i przekazywane dalej do synaps na zakończeniach aksonu.
Przewodzenie informacji w postaci sygnału elektrycznego jest możliwe dzięki temu, że wszystkie neurony są elektrycznie pobudliwe, czyli zdolne do generowania i przewodzenia potencjałów elektrycznych. Niepobudzony neuron utrzymuje potencjał spoczynkowy (będący różnicą między potencjałem elektrycznym wnętrza neuronu a zewnętrznej powierzchni błony) dzięki działaniu leżących w błonie pomp jonowych, które przenoszą określone jony przez błonę i generują różnicę w stężeniu tych jonów po obu stronach błony. Pod wpływem dostatecznie silnego bodźca dochodzi do zmian w przepuszczalności określonych jonów przez błonę, co prowadzi do powstania potencjału czynnościowego - sygnału elektrycznego, który rozprzestrzenia się wzdłuż aksonu do synaps znajdujących się w zakończeniach aksonu.
Neurony powstają w procesie neurogenezy z neuronalnych komórek macierzystych i po zróżnicowaniu nie ulegają już dalszym podziałom komórkowym, neurogeneza zachodzi głównie w okresie prenatalnym (przed narodzeniem), u dorosłych osobników proces ten zachodzi jedynie w określonych częściach mózgu, m.in. hipokampie i opuszce węchowej.
BUDOWA ZEWNĘTRZNA: Neuron składa się z ciała komórki (zwanego perykarionem lub somą) i odchodzących od niego wypustek (zwanych neurytami): dendrytów i aksonów. Cała komórka nerwowa, podobnie jak wszystkie inne komórki zwerzęce, pokryta jest błoną komórkową o grubości ok. 5 nm, która w przypadku komórek nerwowych bywa nazywana neurolemmą.
Rozmiary neuronów: Istnieją ogromne różnice w rozmiarach różnych typów komórek nerwowych u różnych gatunków zwierząt. Ciało komórkowe typowego ludzkiego neuronu ma średnicę ok. 20 mikrometrów. Najmniejsze neurony mają średnicę ok. 5 mikrometrów, największe ok. 120 mikrometrów. Ludzkie aksony mają średnicę w granicach 0,2 do 20 mikrometrów (u bezkręgowców nawet do 1 mm) i długość od kilku mikrometrów do kilku metrów.
BUDOWA WEWNĘTRZNA: Ciało komórki nerwowej (perykarion) zawiera typowe organelle komórki zwierzęcej: jądro komórkowe, gładkie i szorstkie retikulum endoplazmatyczne, cytoszkielet (mikrotubule, mikrofilamenty, a także filamenty pośrednie, które w przypadku neuronów są często nazywane neurofilamentami), mitochondria, aparat Golgiego oraz rybosomy.
Cechą charakterystyczną komórek nerwowych jest występowanie ziarnistości Nissla - gęsto pokrytych rybosomami struktur tworzonych przez szorstkie retikulum endoplazmatyczne. Ich obecność odzwierciedla zdolność neuronów do utrzymywania szybkiego tempa syntezy białek, która zachodzi właśnie w rybosomach.

Skład organelli we wnętrzu dendrytów jest podobny do tego w perykarionie, natomiast we wnętrzu aksonu brak jest szorstkiego retikulum endoplazmatycznego i rybosomów, co oznacza, że wszystkie znajdujące się w nim białka muszę być syntetyzowane w ciele komórki i przetransportowane wzdłuż aksonu. Transport białek z ciała komórki do zakończeń aksonów nazywany jest transportem aksonalnym. Zsyntetyzowane w ciele komórki białka są zamykane w specjalnych pęcherzykach i wędrują wzdłuż leżących w aksonie mikrotubuli dzięki białkom motorycznym zwanym kinezynami, które energię do generowania ruchu pozyskują z ATP. Ruch w przeciwną stronę - od zakończeń aksonu do ciała komórki - odbywa się przy udziale innych białek motorycznych zwanych dyneinami.
SYNAPSA - jest to miejsce komunikacji błony kończącej akson z błoną komórkową drugiej komórki - nerwowej lub komórki efektora (narządu wykonawczego), np. mięśni lub gruczołu. Impuls nerwowy zostaje przeniesiony z jednej komórki na drugą przy udziale substancji o charakterze neuoroprzekaźnika (zwanego czasem neurohormonem) - mediatora synaptycznego (synapsy chemiczne) lub na drodze impulsu elektrycznego (synapsy elektryczne). Synapsy otoczone są przez astrocyty, które uczestniczą w przekazywaniu sygnału między neuronami. Powtarzające się przekazywanie impulsu długotrwale wzmacnia przewodnictwo synaptyczne. Powstawanie sieci neuronów o zwiększonym przewodnictwie jest podstawą uczenia się i zapamiętywania. Nowe synapsy powstają w procesie zwanym synaptogenezą.


*Efapsa, synapsa elektryczna - stanowi ona taki rodzaj synapsy, w której szczelina synaptyczna jest bardzo wąska, co umożliwia bezpośredni kontakt między koneksonami [Konekson to kanał zbudowany z sześciu biaęk zwanych koneksynami, tworzących połączenie szczelinowe, ang. gap junction między dwiema komórkami, umożliwiające przekazywanie informacji za pomocą prądu jonowego. Kanał taki występuje w synapsach elektrycznych. Połączenie szczelinowe tworzone jest przez dwa koneksony różnych komórek ustawione końcami do siebie w poprzek błony komórkowej. Tworzą one przejście, które umożliwia przepływ jonów nieorganicznych i małych cząsteczek (do ok. 1000 Da) rozpuszczalnych w wodzie z cytoplazmy jednej komórki do drugiej. Fragmenty błon komórkowych przy tym połączeniu leżą dokładnie równolegle, bardzo blisko siebie (2-4 nm). Układ ten stanowi elektryczny i metaboliczny łącznik między komórkami] błony presynaptycznej i postsynaptycznej. W takiej synapsie przemieszczanie jonów ładunków elektrycznych i przechodzenie potencjału czynnościowego między błonami odbywa się bez zmiany charakteru impulsu nerwowego. Efapsy mogą pracować dwukierunkowo, a sygnał przekazywany jest przez nie bez opóźnienia.


NEUROGENEZA (ang. neurogenesis) - jest to proces powstawania nowych komórek nerwowych obejmujący zarówno neurogenezę wieku embrionalnego (prenatalnego) jak i neurogenezę wieku dorosłego (postnatalnego). Neurony - główny element tkanki nerwowej swój rodowód wywodzą z komórek płytki nerwowej formowanej z ektodermy we wczesnych fazach rozwoju organizmu. Neurogeneza u ssaków ograniczona jest głównie do okresu prenatalnego i w większości przypadków zachodzi w obrębie przestrzeni okołokomorowej w której powstają neurony formujące korę nową, móżdżek i inne struktury układu nerwowego (Jacobson i in. 1991). Niemniej jednak wiele badań dowodzi, że również w dorosłym móżgu dochodzi do neurogenezy, choć na zdecydowanie niższym poziomie niż w okresie rozwoju zarodkowego (Eriksson PS 1998, Gage FK 1998).
W początkowych etapach formowanie się kory nowej neuroblasty [Neuroblast to komórka macierzysta dla neuronów oraz komórek glejowych. Neuroblasty różnicują się w kierunku neuronów i komórek gleju przede wszystkim w okresie embrionalnym, ale proces neurogenezy zachodzi również w mógowiu ssaków w okresie postnantalnym Wykazano obecność neuroblastów w strefie przyziarnistej zakrętu zębatego i w strefie przykomorowej komór bocznych. Neuroblasty drozofili są modelowymi komórkami w badaniach na neurogenezą u eukariontów. Neuroblasty typu I podlegają asymetrycznym podziałom, samoodnawiając się i tworząc tzw. ganglion mother cells (GMC), z których po kolejnym podziale powstają neurony lub komórki gleju. Neuroblasty typu II również samoodnawiają się, tworząc kolejne neuroblasty i tzw. immature intermediate precursors (INP), które mają zdolność odnawiania swojej puli i tworzenia GMC. GMC mogą podzielić się raz, tworząc dwa niezróżnicowane neurony (postmitotyczne neuroblasty) lub dwie niezróżnicowane komórki gleju (postmitotyczne glioblasty). Z powodu ograniczeń etycznych pozyskiwanie ludzkich neuroblastów jest trudne. Termin neuroblastów wprowadził do neuroanatomii Wilhelm His - niemiecki embriolog, anatom, histolog, wynalazca mikrotomu, w 1889 roku] tworzą warstwę brzeżną która zlokalizowana na brzegu pęcherzyka, zalążka kresomózgowia, zaczyna się rozdzielać. Rezultatem rozwarstwienia jest powstanie podpłytki i zawiązka i warstwy kory mózgowej (warstwy drobinowej). W wyniku procesu zachodzącego zgodnie z regułą inside-out obserwuje się migracje młodych neuronów poprzez wcześniej uformowane, wewnętrzne warstwy neuronów ku powierzchni kory nowej. Neuroblasty tworzące warstwę V (piramidową wewnętrzną) przechodzą przez wcześniej uformowaną warstwę VI (komórek różnokształtnych) by zająć miejsce pod warstwą I. Następnie w ten sam sposób powstaje warstwa IV (ziarnista wewnętrzna) i III (piramidową zewnętrzną) aż w okresie końcowym życia płodowego dochodzi do uformowania warstwy II (ziarnistej zewnętrznej) której komórki migrując przez cztery pierwsze warstwy wypełniają przestrzeń pomiędzy warstwą III i I (Jacobson i in. 1991). Komórkami odgrywającymi najważniejszą role w wędrówce młodych neuronów są komrki Cajala-Retziusa warstwy brzeżnej. Określają one szlak migracji neuroblastów i są źródłem związków ułatwiających młodym neuronom przeciskanie się przez kolejne uformowane warstwy kory nowej (Jacobson i in. 1991).



FUNKCJONOWANIE NEURONÓW: Podstawową funkcją neuronów jest przenoszenie i przetwarzanie informacji w postaci impulsów nerwowych (potencjałów czynnościowych), będących krótkotrwałymi, gwałtownymi zmianami potencjału błony komórkowej neuronu. Impulsy nerwowe w warunkach naturalnych są przewodzone tylko w jednym kierunku [ortodromowo; przewodzenie ortodromowe to sposób przekazywania bodźca w komórce nerwowej polegający na przekazie od ciała komórki (perykarionu) do synaps. Przeciwieństwem jest drugi sposób - przewodzenie antydromowe polegające na transporcie z synaps do ciała komórki - perykarionu] od początkowego segmentu aksonu do synaps znajdujących się na jego zakończeniach. Funkcjonalnie neuron można podzielić na cztery strefy:
POTENCJAŁ CZYNNOŚCIOWY (IGLICOWY) = IMPULS NERWOWY - jest to przejściowa zmiana potencjału błonowego komórki, związana z przekazywaniem informacji. Bodźcem do powstania potencjału czynnościowego jest zmiana potencjału elektrycznego w środowisku zewnętrznym komórki. Wędrujący potencjał czynnościowy nazywany jest impulsem nerwowym. Faza depolaryzacji i repolaryzacji potencjału czynnościowego (iglica) trwa nie więcej niż 1 ms i osiąga maksymalnie wartości około +30 mV. Hiperpolaryzacja [Hiperpolaryzacja to stan nadmiernej polaryzacji błony komórkowej w następstwie zwiększenie elektroujemności wewnątrz komórki; trwa stosunkowo krótko. Błona komórkowa osiąga wówczas potencjał ok. - 80 mV. Podczas hiperpolaryzacji komórka nie jest zdolna do przewodzenia impulsu i przekazywania informacji] następcza może trwać kilkadziesiąt milisekund. W trakcie potencjału czynnościowego neurony stają się niepobudliwe, zaś później, pdoczas hiperpolaryzującego potencjału następczego ich pobudliwość jest zmniejszona. Zjawiska te nazywamy refrakcją bezwzględną i względną. Ze względu na orkes refrakcji bezwzględnej oraz refrakcji względnej komórki nerwowe człowieka nie mogą generować potencjałów czynnościowych z dowolną częstotliwością. Jednak w najbardziej sprzyjających okolicznościach częstotliwość potencjałów czynnościowych może dojść do 100 impulsów na sekundę.
Miejscem powstawania potencjału czynnościowego w komórce nerwowej jest tzw. wzgórek aksonowy, skąd potencjał iglicowy rozprzestrzenia się po powierzchni błony komórkowej wypustki osiowej - aksonu. Generowaniem potencjałów czynnościowych rządzi zasada: "wszystko albo nic": do zapoczątkowania potencjału czynościowego potrzebny jest bodziec o intensywności wystarczającej do zdepolaryzowania neuronu powyżej określonej wartości progowej; wszystkie potencjały czynnościowe w danej komórce osiągają tę samą amplitudę.
PRZEBIEG POTENCJAŁU CZYNNOŚCIOWEGO W KOMÓRCE NERWOWEJ: Jeżeli dokomórkowe prądy kationów przeważają nad odkomórkowymi, następuje depolaryzacja od poziomu potencjału spoczynkowego (ok. -70 mV) do potencjału krytycznego czyli progowego (ok. -50 mV). Po osiągnięciu potencjału progowego, następuje otwarcie bramkowanych elektrycznie (zależne od napięcia) kanałów przewodzących odkomórkowo kationy potasowe oraz kanałó przewodzących kationy sodowe do wnętrza komórki. Wskutek różnicy stężeń i potencjałów kationy sodowe szybko napływają do wnętrza komórki, niwelując różnicę potencjałów pomiędzy środowiskiem zewnątrz- i wewnątrzkomórkowym do zera (depolaryzują błonę komórkową). Jest to początek tzw. potencjału iglicowego. Ze względu na dużą początkową różnicę stężeń, mimo wyrównania potencjałów, kationy sodowe napływają w dalszym ciągu do wnętrza komórki, powodując odwrotną polaryzację błony, tzw. nadstrzał dochodzący do +35 mV. Następuje to stosunkowo szybko; w ciągu ok 2 ms następuje już całkowita inaktywacja kanałów sodowych. Po rozpoczęciu aktywacji sodowej, w odpowiedzi na depolaryzację, następuje aktywacja potasowa, czyli otwarcie kanałów dla odkomórkowego prądu kationów potasowych. Powoduje to zmianę potencjału wnętrza komórki z powrotem na ujemny, czyli repolaryzację. W tym czasie nie jest możliwe ponowne pobudzenie komórki, jest to okres refrakcji bewzględnej [reflaksja to właściwość komórek pobudliwych, a także błon komórkowych wypustek tych komórek, polegająca na okresowej niewrażliwości na stymulujące je bodźce po przejściu potencjału czynnościowego: refkracja bezwzględna to stan, w którym komórka nie jest w stanie odpowiedzieć na żaden bodziec - tak jest zaraz po przejściu potencjału, a z czasem przechodzi w --> refrakcję względną - gdy bodziec o większym niż fizjologicznie nasileniu może wywołać odpowiedź komórki]. W czasie trwania potencjału iglicowego, a zwłaszcza pod koniec - proporcje kationów po obu stronach błony są odwrócone (Na+ wewnątrz, K+ na zewnątrz komórki). Przez cały czas trwania potencjału czynnościowego, aktywna jest pompa sodowo-potasowa oraz inne układy transportujące jony. Poziom aktywności pompy sodowo-potasowej jest bowiem uzależniony m.in. od stężenia kationów sodowych we wnętrzu komórki. Zatem w czasie trwania potencjału iglicowego pompa działa z najwyższą możliwą prędkością. W ciągu kolejnych kilkudziesięciu milisekund przywraca proporcje kationów sodowych i potasowych po obu stronach błony komókowej. Jednak początkowo błona jest jeszcze w stanie hiperpolaryzacji - różnica potencjałów przekracza wartość spoczynkową. Jest to okres refrakcji względnej, kiedy komórkę da się pobudzić, ale dużo większym bodźcem - ze względu na większą "odległość" od poziomu potencjału krytycznego. W pewnym stopniu, w różnych tkankach, w opisanych procesach biorą udział również inne jony, głównie jony chlorkowe (Cl-) oraz wapniowe (Ca2+).
ZASADA WSZYSTKO ALBO NIC: Przewodzenie sygnałów przez neuron podlega zasadzie wszystko albo nic, tzn. neuron wytwarza potencjał czynnościowy lub nie. Wszystkie powstające potencjały czynnościowe w danej komórce nerwowej mają tę samą wielkość, bez względu na wielkość bodźca (o ile tylko jest on wystarczająco intensywny, aby wywołać powstanie potencjału). Silniejsza stymulacja neuronów (bodźcami ponadprogowymi) nie prowadzi do wytwarzania silniejszych potencjałów. Może natomiast prowadzić do zwiększenia częstotliwości wytwarzania potencjałów przez neuron.
HISTORIA BADANIA FUNKCJI NEURONÓW: Znaczna część pierwotnej wiedzy o aktywności elektrycznej neuronów pochodzi z eksperymentów na aksonach wielkich kałamarnic. W 1937 roku John Zachary Young zaproponował aksony kałamarnic jako model użyteczny do studiów nad elektrycznymi właściwościami neuronów. Są one dużo większe od ludzkich neuronów, więc lepiej nadawały się do eksperymentów w tamtych czasach.

- The Squid and its giant nerve fibre - Part 1
W mózgu człowieka jest ponad 10000 specyficznych typów neuronów, ale ogólnie wyróżniamy trzy główne typy neuronów ze względu na pełnione na nie funkcje: neurony ruchowe (przekazują informacje motoryczne), neurony czuciowe (przekazują informacje czuciowe) i interneurony (przekazują informacje różnych typów komórek nerwowych). Neurony mogą także przyjmować różne kształty i rozmiary (zobacz to na poniższych rysunkach wykonanych przez Cajala - hiszpańskiego histologa, neuroanatoma, prekursora neurobiologii, który wraz z Camillo Golgim nagrodzony został Nagrodą Nobla w dziedzinie medycyny w roku 1906).
 FUNKCJOWANIE SYNAPS CHEMICZNYCH I ELEKTRYCZNYCH (ANIMACJA): KLIK!
FUNKCJOWANIE SYNAPS CHEMICZNYCH I ELEKTRYCZNYCH (ANIMACJA): KLIK!




 NEURONY LUSTRZANE:
NEURONY LUSTRZANE:

LICZBA NEURONÓW: Szacuje się, że ludzki mózg zawiera ok. 1,5-1,6 x 1011 neuronów i 1014 synaps. Ogromna większość neuronów znajduje się w móżdżku, a najliczniejszą populacją komórek są małe neurony ziarniste móżdżku.
Osiągnięcia Hiszpańśkiej Szkoły Neurologicznej: Do czasu odkrycia Ramona y Cajala i Camillo Golgiego uważano, że mózg jest jednolitą masą, ponieważ metodami, jakimi dysponowano i stosowano, nie potrafiono uzyskać obrazu komórek nerwowych.
Obaj naukowcy zastosowali tę samą metodę barwienia preparatów histologicznych, która oparta była na nowej wówczas technice fotograficznej. Została ona opracowana w roku 1873 przez Golgiego (tzw. metoda Golgiego). Golgi uważał, że struktura obserwowanych tkanek ejst siecią wielu rozgałęziających, bezpośrednio połączonych nerwowych kanalików.
Ramon y Cajal - zafascynowany wyglądem zabarwionych preparatów Golgiego, które zobaczył po kilkunastu latach (w roku 1887) - przystąpił do analogicznych eksperymentów. Intensywne badania objęły liczne preparaty, pobrane z tkanki pochodzącej od ludzi i wielu różnych zwierząt i ich płodów (króliki, psy, świnki morskie, kurczęta, myszy, szczury, żaby, ryby). Kształty zabarwionych neuronów precyzyjnie rysował. Umożliwiło mu to sformułowanie "doktryny neuronów" - teorii, że aksony poszczególnych neuronów nie łączą się między sobą, lecz każdy akson kończy się zawsze na dendrycie sąsiedniego neuronu (protoplazma obu komórek nie miesza się ze sobą, stąd pomiędzy nimi istnieje przestrzeń zwana synapsą). Odkrycie to zrewolucjonizowało badania struktury i zasad działania układu nerwowego.
W 2017 roku archiwum Santiago Ramona y Cayala i jego współpracowników zostało wpisane na listę Pamięci Świata jako Archives of Santiago Ramon Y Cajal and the Spanish Neurohistological School. Archiwum, zawierające prace uczonego (preparaty, rysunki, fotografie, opisy) oraz wytworzone przez niego dzieła sztuki (rysunki i obrazy), fotografie, jego korespondencję, notatki, książki (napisane oraz będące w jego bibliotece) są ważnym źródłem do poznania historii rozwoju neurobiologii.
Więcej informacji na temat Ramona Cajala znajdziesz: tutaj.
Hiszpańska szkoła neurologiczna:
Niezwykłą cechą kariery naukowej Ramona Cajala była zdolność przyciągnięcia wielu młodych współpracowników, którzy z czasem stali się mistrzami w swoich dziedzinach badawczych, przyczyniając się do fundamentalnych odkrycia do naszej obecnej koncepcji neuronauki: Pío del Río-Hortega (Portillo -Valladolid - 1882-1945) zidentyfikował dwa z czterech głównych typów komórek nerwowych: oligodendrogleju i mikrogleju; Fernando de Castro (Madryt, 1896-1967) zidentyfikował dokładne położenie chemoreceptorów tętniczych w ciele szyjnym, fundamentalne dla odruchów sercowo-oddechowych; Rafael Lorente de Nó (Saragossa, 1902-1990) stworzył wiele dominujących koncepcji dotyczących funkcji kory mózgowej; Nicolás Achúcarro (Bilbao, 1880-1918), jeden z pierwszych uczniów Cajala, był entuzjastą neuropatologii, który wniósł w duży wkład w badania nad procesami neurodegeneracji (przez pewien czas pracował z Alois Alzheimerem); Jorge Francisco Tello (Alhama de Aragón -Zaragoza-, 1880-1958) przeprowadził badania, które zostały odtąd odremontowane przez nasz obecny pogląd na regenerację aksonów i nerwów; Pedro Ramón y Cajal (Larrés-Huesca-, 1854-1950), brat Santiago, specjalizujący się w układach nerwowych owadów i bezkręgowców. Najmłodsi członkowie tej szkoły, De Castro i Lorente de Nó, przeszli od czysto histologicznej do bardziej fizjologicznej orientacji swoich badań.
Łącznie ta grupa hiszpańskich neuronaukowców skupionych przez Santiago Ramona y Cajala jest zbiorowo znana jako hiszpańska szkoła neurologiczna (lub neurologiczna), lub po prostu Szkoła Cajal i stanowi jeden z najbardziej wyjątkowych przykładów szkół naukowych w historii nauki z powodu jej licznych naukowych sukcesów i odkryć.
El Archivo de Santiago Ramón y Cajal y la Escuela Española de Neurohistología (Archives of Santiago Ramon Y Cajal and the Spanish Neurohistological School) – to zbiory książek, dokumentów i przedmiotów pozostałych po Santiago Ramonie y Cajalu - hiszpańskim pionierze neurologii oraz kilku jego uczniach i współpracownikach. W skład wchodzą książki, rękopisy, preparaty anatomiczne (głównie neurologiczne), rysunki i obrazy (zarówno ilustracje medyczne jak i dzieła artystyczne), korespondencja, zdjęcia (zarówno naukowe, jak i inne) oraz rzeczy używane przez naukowców.

--> Archive del Rio-Hortega - pozostałe po Pio del Rio-Hortega, który w czasie wojny 1936-1939 wyemigrował i zmarł w Argentynie. Część zbiorów została przewieziona na uniwersytet w Valladolid i jest dostępna publicznie, druga część jest w posiadaniu rodziny. Całość obejmuje 35 publikacji naukowych, 5 rękopisów (jedna książka i 4 materiały konferencyjne), 750 listów i kartek pocztowych, 50 dokumentów (dyplomy, programy itp), 11 rysunków naukowych, 60 preparatów oraz zdjęcia. Pio del Rio-Hortega był hiszpańskim lekarzem neurologiem, neuroanatomem i neuropatologiem, jednym z czołowych przedstawicieli hiszpańskiej szkoły neurologicznej.
--> Archive Fernando de Castro - w posiadaniu rodziny Fernando de Castro znajdują się: 3 książki, 2 podręczniki i 65 publikacji (w tym 9 dotyczących Cajala oraz Szkoły), rękopis książki napisanej w 1933 wspólnie z Cajalem, 11 innych dokumentów i około 2 tysięcy listów. Oprócz tego 609 rysunków (w tym 531 sporządzonych przez Castro), 4129 preparatów i tysiące fotografii różnych autorów. Przechowywane są też 3 mikroskopy i inne pamiątki.
RYSUNKI FERNANDO DE CASTRO ZNAJDZIESZ: TUTAJ.
Zaraz po uzyskaniu doktoratu pracujący nad strukturą zwojów czuciowych i wegetatywnych, wraz z umiejętnościami nabytymi w tym czasie w zakresie technik histologicznych, Fernando de Castro zaczął badać unerwienie rejonu tętniczo-szyjnego. W pierwszych dwóch dekadach XX wieku rejon ten był był celem licznych badań, po pojawieniu się opisu odruchów oddechowych Heinricha Heringa. Dwa istotne badania De Castro stworzyły anatomiczne podstawy dla wyjaśnienia istoty owych odruchów (badania publikowane w 1926 i 1928 roku). W nich Fernando de Castro opisał rozdzieloną dystrybucję baroreceptorów w zatoce tętnicy szyjnej, a także chemoreceptory w ciele szyjnym (glomus caroticum). Wykazał również sensoryczną naturę tego unerwienia, oddzielonego od włókien nerwowych pochodzących z wegetatywnych zwojów. Najbardziej znacząca fizjofarmakologiczna grupa owych czasów, związana z osobą Corneille'a Heymansa (1892-1968) z Ghent University (Belgia) przedstawiła znaczenie pracy De Castro i, po zaproszeniu Fernanda de Castro do Ghent, aby wspólnie przeprowadzili różne eksperymenty, szkoła Ghent School postanowił skoncentrować się na carotid body (ciele szyjnym) w celu zbadania chemoreceptorów tętniczych. Ta decyzja była kluczowa dla ówczesnej pracy Corneille Heymansa i to on otrzymał Nagrodę Nobla z 1938 roku w dziedzinie fizjologii lub medycyny, podczas gdy Fernando de Castro znajdował się pod ciężkim zagrożeniem życia w Madrycie, broniąc Instytutu Cajal przed grabieżą podczas hiszpańskiej wojny domowej (1936-1939). Królewska Szwedzka Akademia Nauk (The Swedish Nobel Academy) zapomniała o opisie De Castro, chociaż już wielokrotnie był on przywoływany na świecie i zostął uznany za fundamentalny wkład młodego neurologa z Hiszpanii i jego zasług; pojawiały się głosy, by podzielić Nagrodę Nobla pomiędzy Heymansem i de Castro.
Fernando de Castro kontynuował swoje badania na ten temat. Ważnymi publikacjami były te opublikowane w 1940, 1944, 1950, 1951, 1962 i pośmiertelnie w 1968.




Rys: Typowe waskularne zakończenia astrocytów w opuszce węchowej (1916):

--> Archive Pedro Ramon y Cajal - archiwum pozostawione przez Pedro Ramona y Cajala, brata Santiago. Przechowywane przez krewnych Pedro. Składa się z 20 publikacji naukowych, 2 manuskryptów Santiago i 8 listów oraz manuskryptów Pedro, 50 rysunków oraz 150 preparatów. W skład kolekcji wchodzi też 350 płyt ze zdjęciami stereoskopowymi (fotografia stereoskopowa skłąda się z dwóch zdjęć tego samego obiektu, ale wykonanych z różnych punktów widzenia. Patrzący przez stereoskop widz ma wrażenie przestrzenności i trójwymiarowości oglądanej sceny) wykonanymi przez Santiago.
--> Archive Lorente de No - po Rafaelu Lorente de No pozostało 65 rysunków i 100 preparatów, które są przechowywane przez Jorge Larriva-Sadh, jednego z uczniów Rafaela Lorente de No. W latach 30-tych Lorente de No opublikował cenione prace dotyczące anatomii kory mózgowej, dróg słuchowych i formacji hipokampa. Był pierwszym, który opisał opóźnienie synaptyczne.
PRZYKŁADOWE RYSUNKI CAJALA:
Źródła: nobelprize.org
Rys.01.
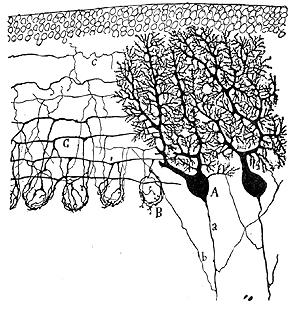
Aksony komórek Purkiniego to jedyne włókna, jakie opuszczają korę móżdżku. Natomiast do kory móżdżku docierają dwa typy włókien - pnące (rozpoczynające się w jądrze oliwki) i kiciaste (inaczej mszyste; rozpoczynające się w rdzeniu kręgowym i neuronach jąder mostu). Włókna pnące kończą się na dendrytach komórek Purkiniego, w warstwie drobinowej. Pojedynce włókno pnące pobudza zaledwie kilka neuronów Purkiniego, ale jest to pobudzenie o niezwykłej sile. Połączenia włókien pnących z komórkami Purkiniego to jedne z najsilniejszych synaps w układzie nerwowym, a każde wyładowanie wywołuje pobudzający potencjał postsynaptyczny. Wyładowania te są rzadkie - 1/s. Wyładowania z włókien pnących nakładają się na pobudzenia pochodzące z włókien kiciastych, modyfikując ich oddziaływanie na komórki Purkiniego. Ogólnie, włókna pnące dostarczają komórkom Purkiniego informacji dotyczących wzorca ruchowego wykonywanej czynności. Włókno kiciaste pobudza wiele komórek ziarnistych, których aksony po rozdwojeniu tworzą włókna równoległe. Włókna równoległe są niezmiernie liczne i tworzą około 200 000 synaps na każdej komórce Purkiniego. Dzięki częstym wyładowaniom następuje sumowanie czasowe i przestrzenne, które powoduje stałe pobudzenie komórek Purkiniego. Włókna kiciaste (za pośrednictwem włókien równoległych) są dla komórek Purkiniego źródłem informacji o "poleceniach", jakie zostały wydane motoneuronom rogów przednich rdzenia przez korę mózgu oraz o stanie realizacji tych "poleceń" przez mięśnie.
Dendryty apikalne komórek Purkinjego mają ogromną liczbę kolców dendrytycznych (ponad 80 000 na komórkę).
Komórki Purkinjego zawierają kwas gamma-aminomasłowy (GABA) - neurotransmiter hamujący (przez co działają hamująco na swoje miejsca docelowe). Charakterystyczną cechą tych komórek jest także ekspresja białka kalbindyny. Barwienie kalbindyny z mózgu szczura po przewlekłym jednostronnym uszkodzeniu nerwu kulszowego sugeruje, że nowe neurony Purkniego mogą być tworzone w dorosłym mózgu, inicjując organizowanie nowych płatów w móżdżku.

Rys.02.
 Rys.03.
Rys.03.
Na rysunku tym możemy zaobserwować różnorodność form morfologicznych neuronów - widzimy np. tutaj neurony dwubiegunowe.

 Rys.05.
Rys.05.


HISTORIA: Pierwszymi poważnymi badaniami nad opisem gleju zajął się Santiago Ramon y Cajal (1852-1934) który scharakteryzował trzy podstawowe rodzaje komórek glejowych. Doniósł on o obecności komórek o kształcie gwieździstym - nieświadom tego że są to astrocyty, komórek włóknistych z licznymi wypustkami i komórek protoplazmatycznych występujących licznie w istocie szarej. Jak się okazało później jest to mikroglej. Niezależnie odkryty przez Nissla i Robertsona, dokładniejszy opis zyskał jednak dzięki uczniowi Cajala, Pio del Rio-Hortedze (1882-1945). W wyniku barwienia solami srebra Rio-Hortega wyróżnił specyficzny rodzaj komórek zawierających jądro o wyraźnie wydłużonym kształcie. Scharakteryzowane przez niego komórki mikrogleju zyskały od tej pory miana komórek Hortegi. Następny "krok milowy" w badaniach nad mikroglejem należał znów do Hortegi, który opisywał rolę odkrytego mikrogleju w patologii centralnego układu nerwowego. W późniejszych latach, w wyniku zidentyfikowania komórkowo specyficznych markerów narastały kontrowersje wokół rozwoju embrionalnego jak i pochodzenia mikrogleju. Te i inne wątpliwości rozwiały zaawansowane badania w latach 80. z użyciem markerów lektynowych oraz rozwiniętych technik immunocytochemicznych.
Zgodnie z mezodermalną teorią pochodzenia komórek mikrogleju monocyty infiltrują parenchymę mózgu we wczesnych okresach życia embrionalnego jako mikroglej amebowaty. Zaznaczyć trzeba że napływ mikrogleju do mózgu jest dodatnio skorelowany z rozwojem unaczynienia tkanki nerwowej co również przemawia za mezodermalnym źródłem mikrogleju (Perry i in. 1985, Miyake i in. 1984, Hurley i in. 1996). Opuszczenie naczyń krwionośnych przez te komórki jest możliwe gdyż bariera krew-mózg tworzona w tym czasie przez astrocyty nie jest jeszcze dokładnie zamknięta w tym stadium życia organizmu. Po przeniknięciu dochodzi do transformacji mikrogleju amebowatego w mikroglej spoczynkowy charakteryzujący się obkurczoną częścią cytoplazmatyczną oraz licznymi rozgałęzieniami (Boya i in. 1991, Fedorolf i in. 1995).
Jako alternatywne źródło mikrogleju podaje się neuroektodermę, z której wywodzą się glioblasty będące prekursorami astrocytów i oligodendrocytów (Fujita i in. 1975, 1980). Jednyzm z dowodów ppopierających teorię ektodermalnego pochodzenia mikrogleju jest fakt, iż mikroglej, jak i komórki progenitorowe 0-2A, z których mogą się rozwinąć oligodendrocyty oraz astrocyty typu drugiego, posiadają jednakowe właściwości histochemiczne. Poza tym zidentyfikowano homologiczne epitopy powierzchniowe zlokalizowane na astrocytach, jak i na mikrogleju, a rozpoznawane przez przeciwciała monoklonalne LN-1 (Dickson i in. 1989). Zgodnie z neuroektodermalną teorią o pochodzeniu mikrogleju zakłada się, że mikroglej wykształcił się jako samodzielna linia z puli komórek macierzystych neurogleju. Faktem przemawiającym za tą teorią jest to, że glioblasty ze strefy okołokomorowej wykształconej z cewki nerwowej są prekursorami komórek Hortegi. Znaczyłoby to, że oligodendrocyty astrocyty i mikroglej są grupami siostrzanymi pochodzącymi z tej samej linii multipotencjalnych komórek neuroektodermalnych (Fujuta i in. 1975, 1980).
Ze względu na brak jednoznacznych i niepodważalnych faktów przemawiających za jedną z opcji pochodzenia mikrogleju proponowane jest alternatywne heterogeniczne źródło (De Groot i in.). Według tego złożenia część mikrogleju zasiedlającego tkankę nerwową jest pochodzenia mezodermalnego, a część neuroektodermalnego. Bez względu na pochodzenie mikrogleju, przyjmuje on w mózgu formy i funkcje charakterystyczne tylko dla tego rodzaju komórek gleju w dużym stopniu zależne od stanu fizjologicznego tkanki.
Funkcja mikrogleju: Mikroglej stanowiący około 5-20% populacji komórek nieneuronalnych w mózgu występuje w warunkach normalnych w formie spoczynkowej. Główne funkcje fizjologiczne mikrogleju sprowadzają się do monitorowania mikrośrodowiska tkanki, usuwania umierających neuronów i reakcji w wyniku stwierdzenia obecności obcego antygenu (Nakajima i in. 1993). W formie nieaktywnej mikroglej posiada liczne i długie rozgałęzienia części cytoplazmatycznej komórki. Ta specyficzna struktura prawdopodobnie ułatwia potencjalną reakcję tych komórek jako specyficznego dla tkanki nerwowej elementu układu immunologicznego.
Charakterystyczna transformacja mikrogleju z formy spoczynkowej do aktywowanej została poznana i opisana przez Rio Hortege prawie wiek temu. W wyniku aktywacji dochodzi do przemian morfologicznych którym towarzyszy: zwiększenie rozmiaru ciała komórki oraz obkurczenie rozgałęzień. Na poziomie molekularnym dochodzi do ekspresji białek adhezyjnych, reorganizacji cytoszkieletu oraz ekspresji elementów kompleksu zgodności tkankowej MHC typu I jak i II (Perry i in. 1987, 1989, Aloisi i in. 2000).
Zaobserwowano, że aktywowanie mikrogleju może odgrywać rolę w ciężkich postaciach zaburzeń psychiatrycznych.
MIKROGLEJ W USZKODZONEJ TKANCE NERWOWEJ: W wyniku uszkodzenia tkanki nerwowej dochodzi w mózgu do odpowiedzi komórkowej i chemicznej otaczających komórek które to reakcje są efektem fizycznego naruszenia integralności tkanki nerwowej, jak i zmian w lokalnym mikrośrodowisku, np. zmiany homeostazy jonowej. Jednocześnie z uszkodzeniem uruchamianych jest wiele procesów mających na celu doprowadzić z jednej strony do ochrony przed czynnikami uszkadzającymi, a z drugiej do naprawy powstałych zniszczeń tkanki (Berkenbosch i in. 1992).
Odpowiedzi komórkowej na czynnik uszkadzający w większości przypadków towarzyszy: proliferacja i migracja komórek mikrogleju i astrocytów, produkcja cytokin prozapalnych, funkcjonalne zmiany w śródbłonku naczyń krwionośnych oraz rekrutacja z krwiobiegu komórek układu immunologicznego w obręb uszkodzonej tkanki (Dong i in. 2001, Aloisi i in. 2001). W uszkodzonych neuronach dochodzi do ekspresji wczesnych genów odpowiedzialnych za stymulację i aktywowanie astrocytów i mikrogleju (Neumann i in. 2001, Raivich i in. 1999). A poprzez przerwaną ciągłość bariery krew-mózg aktywowane zostają monocyty z krwi obwodowej które na wzór mikrogleju ulegają następnie transformacji morfologicznej (Fujuta i in. 1998, Leong i in. 1992, Maxwell i in. 1990).
Aktywacja mikrogleju jaką wywołuje uszkodzenie tkanki związana jest ze wzrostem poziomu czynników o funkcji immunologicznej. W okresie tym może dojść do indukcji ekspresji szerokiej rzeszy receptorów powierzchniowych które przyspieszają odpowiedź immunologiczną. Pośród nich są receptory biorące udział w rozpoznawaniu cząsteczek związanych z patogenem, receptory komplementu (np. CR1, CR3, CR4), receptory cytokin (np. TNFRI, TNFRII, IL-1RI, IL-12R) oraz chemokin (np. CCR2, CCR3, CXCR4, CX3CR1) jak i receptory ułatwiające interakcje z układem immunologicznym jak np. z limfocytami T czy immunoglobulinami (np. Fc RI, RII, RIII) (Aloisi i in. 2001, Perry i in. 1992, Nakajima i in. 1993).




b) Oligodendrocyty - są to komórki gleju formujące osłonki mielinowe w centralnym układzie nerwowym. Nazwa oligodendrocytów została po raz pierwszy wprowadzona przez Pio del Rio-Hortegę (1882-1954). Oligodendrocyty znane są głównie ze względu na zdolność formowania osłonek mielinowych w ośrodkowym układzie nerwowym. W procesie mielinizacji liczne wypustki oligodendrocytów w wyniku kontaktu z aksonem zaczynają obwijać go swoją błoną komórkową w postaci spiralnych zwojów, tworząc w ten sposób osłonkę mielinową. Jeden ilogodendrocyt osłania fragmenty aksonów kilku komórek, w przeciwieństwie do lemocytu (odpowiednika oligodendrocytu w obwodowym układzie nerwowym), który na ogół wytwarza osłonkę mielinową dookoła jednego włókna nerwowego. Liczba osłonek tworzonych przez jedną komórkę oligodendrocytu waha się, w zależności od rejonu centralnego układu nerwowego, od 40 w nerwie wzrokowym szczura do pojedynczej osłonki na mocno zmielinizowanych szlakach rdzeniowych w rdzeniu kręgowym kota.
Podczas rozwoju rdzenia kręgowego, komórki prekursorowe dla oligodendrocytów (ang. oligodendrocyte precursor cells - OPCs) wywodzą się głównie z brzusznej (ang. ventral) części cewki nerwowej (ang. neural tube). W wyniku antagonistycznego działania czynników tj. Shh (ang. sonic hedgehog) oraz wybranych białek z rodziny BMP (ang. bone morphogenic proteins) dochodzi do różnicowania się oligodendrocytów, które następnie kolonizują białą substancję (ang. white matter). Dodatkowo do brzusznej populacji oligodendrocytów, powstaje grupa oligodendrocytów generowanych w centralnej części oraz grzbietowej stronie rdzenia kręgowego. Komórki prekursorowe dla oligodendrocytów pozostaję rozsiane w centralnym układzie nerwowym stanowiąc pulę komórek uczestniczącą w odnawianiu pulu dorosłych oligodendrocytów. Licznie prezentowane w okresie postmitotycznym w centralnym układzie nerwowym, oznaczane poprzez ekspresje specyficznego markera - NG2 - przez wielu badaczy brane są za osobną grupę gleju, zwaną polidendrocytami.
W wyniku uszkodzenia centralnego układu nerwowego, jak również w wyniku chorób neurodegeneracyjnych, dochodzi także do degeneracji i śmierci oligodendrocytów.
Oligodendrocyty eksprymują na swojej powierzchni serie charakterystycznych białek - Nogo-A, MAG, OMgp i inne, które wiązane są silnie z ich hamującymi właściwościami na wzrost zakończeń nerwowych po uszkodzeniu. Teza ta jednak stoi przed wyzwaniem rzuconym przez grupę prof. Schwaba pokazującą nagły i silny wzrost zakończeń nerwowych wzdłuż zmielinizowanych szlaków nerwowych nie tworzących rozgałęzień do sąsiadujących szlaków. Nazwano to z angielskiego guard rai poprzez negatywną stymulację. Co więcej, ostatnie badania in vivo sugeruję, że MAG, komponent mieliny uznawany za inhibitor wzrostu, w rzeczywistości wzmaga wzrost zakończeń nerwowych.
c) Komórki ependymalne (Wyściółka) - łac. ependyma - jest to jednowarstwowy nabłoneks ześcienny wyściełający jamy ośrodkowego układu nerwowego: komór mózgu i wodociągu mózgu, a także kanału środkowego rdzenia kręgowego. Wywodzi się z nabłonka cewy nerwowej. Komórki wyściółki, inaczej ependymocyty, są typem komórek glejowych, posiadają liczne mikrokosmki na powierzchni zwróconej do światła komór. Na powierzchni podstawnej ependymocytów znajdują się liczne wgłobienia i wypustki, za pomocą których komórki te łączą się z astrocytami.
Funkcją ependymy wchodzącej w skład bariery krew-mózg jest uczestniczenie w wymianie substancji pomiędzy płynem mózgowo-rdzeniowym a tkanką nerwową.
d) Komórka Schwanna, także lemocyt, lemmocyt - komórka glejowa obwodowego układu nerwowego występująca w zespołach.

e) Komórki satelitarne (amficyty) - są to komórki glejowe zlokalizowane w obwodowym układzie nerwowym. Przylegają do perikarionów neuronów zwojów nerwów czaszkowych, rdzeniowych i współczulnych. Posiadają spłaszczone, owalne jądra komórkowe. Pochodzą z grzebienia nerwowego. W zwojach są spłaszczone i przylegają do ciał komórek nerwowych. Ich znaczenie czynnościowe nie jest w pełni jasne.
Tkanka ta złożona jest z licznych komórek nerwowych - neuronów. Składają się one z ciała komórki zawierającego jądro. Od tego ciała odchodzą krótkie wypustki zwane dendrytami oraz najczęściej jedna, długa i rozgałęziona na końcu wypustka - neuryt. Mogą go otaczać osłonki mielinowe. Dendryty odbierają bodźce i przekazują je do ciała komórki nerwowej, a stąd przez neuryt informacja trafia do następnej komórki nerwowej. Dzięki dendrytom i neurytom komórki nerwowe mogą spełniać swoje funkcje, czyli odbierać i przekazywać bodźce ze środowiska zewnętrznego i wewnętrznego do centralnego układu nerwowego - mózgu i rdzenia kręgowego. Centralny układ nerwowy, od którego odchodzą liczne włókna nerwowe pełni nadrzędną funkcję w stosunku do innych układów i całego organizmu. Układ nerwowy scala, kontroluje wszystkie czynności życiowe i funkcjonowanie żywego organizmu. Z tkanki nerwowego zbudowane są: ośrodkowy układ nerwowy: mózg i rdzeń kręgowy i obwodowy układ nerwowy.
KOMÓRKI NERWOWE:
NEURON (Komórka nerwowa) - to rodzaj elektrycznie pobudliwej komórki zdolnej do przetwarzania i przewodzenia informacji w postaci sygnału elektrycznego. Neurony są podstawowym elementem układu nerwowego zwierząt. Najwięcej neuronów znajduje się w ośrodkowym układzie nerwowym w skład którego wchodzi mózgowie oraz rdzeń kręgowy.
 |
| Źródło: docplayer.pl - "Tkanka nerwowa, centralny układ nerwowy" - prezentacja użytkownika Bartłomiej Mróz. |
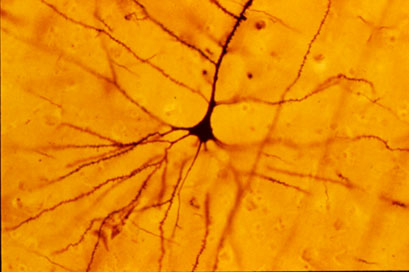 |
| Źródło: Wikipedia. Neuron piramidowy z kory mózgowej człowieka (barwiony metodą Golgiego). |
Przewodzenie informacji w postaci sygnału elektrycznego jest możliwe dzięki temu, że wszystkie neurony są elektrycznie pobudliwe, czyli zdolne do generowania i przewodzenia potencjałów elektrycznych. Niepobudzony neuron utrzymuje potencjał spoczynkowy (będący różnicą między potencjałem elektrycznym wnętrza neuronu a zewnętrznej powierzchni błony) dzięki działaniu leżących w błonie pomp jonowych, które przenoszą określone jony przez błonę i generują różnicę w stężeniu tych jonów po obu stronach błony. Pod wpływem dostatecznie silnego bodźca dochodzi do zmian w przepuszczalności określonych jonów przez błonę, co prowadzi do powstania potencjału czynnościowego - sygnału elektrycznego, który rozprzestrzenia się wzdłuż aksonu do synaps znajdujących się w zakończeniach aksonu.
 |
| Źródło: Wikipedia. Schemat budowy neuronu. |
BUDOWA ZEWNĘTRZNA: Neuron składa się z ciała komórki (zwanego perykarionem lub somą) i odchodzących od niego wypustek (zwanych neurytami): dendrytów i aksonów. Cała komórka nerwowa, podobnie jak wszystkie inne komórki zwerzęce, pokryta jest błoną komórkową o grubości ok. 5 nm, która w przypadku komórek nerwowych bywa nazywana neurolemmą.
Rozmiary neuronów: Istnieją ogromne różnice w rozmiarach różnych typów komórek nerwowych u różnych gatunków zwierząt. Ciało komórkowe typowego ludzkiego neuronu ma średnicę ok. 20 mikrometrów. Najmniejsze neurony mają średnicę ok. 5 mikrometrów, największe ok. 120 mikrometrów. Ludzkie aksony mają średnicę w granicach 0,2 do 20 mikrometrów (u bezkręgowców nawet do 1 mm) i długość od kilku mikrometrów do kilku metrów.
BUDOWA WEWNĘTRZNA: Ciało komórki nerwowej (perykarion) zawiera typowe organelle komórki zwierzęcej: jądro komórkowe, gładkie i szorstkie retikulum endoplazmatyczne, cytoszkielet (mikrotubule, mikrofilamenty, a także filamenty pośrednie, które w przypadku neuronów są często nazywane neurofilamentami), mitochondria, aparat Golgiego oraz rybosomy.
Cechą charakterystyczną komórek nerwowych jest występowanie ziarnistości Nissla - gęsto pokrytych rybosomami struktur tworzonych przez szorstkie retikulum endoplazmatyczne. Ich obecność odzwierciedla zdolność neuronów do utrzymywania szybkiego tempa syntezy białek, która zachodzi właśnie w rybosomach.
- Tigroid (ciało Nissla, ziarenka Nissla, ziarnistości Nissla) - łac. Globule substantiae basophilicae - są to grudki substancji zasadochłonnej w komórkach nerwowych (neuronach) znajdowane w ciałach komórek nerwowych i dendrytach. Tigroid jest specyficzną dla neuronów postacią szorstkiej siateczki śródplazmatycznej i miejscem intensywnej syntezy białek (jest skupieniem rybosomów oraz RNA). Obecność tigroidu jest jedną z przyczyn szarej barwy skupisk ciał komórek nerwowych - stąd noszą one nazwę istoty szarej. Zostały opisane w 1894 roku przez Franza Nissla. Nazwę "tigroid" należy z kolei kojarzyć z tygrysem (Panthera tigris) - z powodu wyglądu tychże grudek.

Skład organelli we wnętrzu dendrytów jest podobny do tego w perykarionie, natomiast we wnętrzu aksonu brak jest szorstkiego retikulum endoplazmatycznego i rybosomów, co oznacza, że wszystkie znajdujące się w nim białka muszę być syntetyzowane w ciele komórki i przetransportowane wzdłuż aksonu. Transport białek z ciała komórki do zakończeń aksonów nazywany jest transportem aksonalnym. Zsyntetyzowane w ciele komórki białka są zamykane w specjalnych pęcherzykach i wędrują wzdłuż leżących w aksonie mikrotubuli dzięki białkom motorycznym zwanym kinezynami, które energię do generowania ruchu pozyskują z ATP. Ruch w przeciwną stronę - od zakończeń aksonu do ciała komórki - odbywa się przy udziale innych białek motorycznych zwanych dyneinami.
 |
| Źródło: Wikipedia. Połączenie synaptyczne. |
SYNAPSA - jest to miejsce komunikacji błony kończącej akson z błoną komórkową drugiej komórki - nerwowej lub komórki efektora (narządu wykonawczego), np. mięśni lub gruczołu. Impuls nerwowy zostaje przeniesiony z jednej komórki na drugą przy udziale substancji o charakterze neuoroprzekaźnika (zwanego czasem neurohormonem) - mediatora synaptycznego (synapsy chemiczne) lub na drodze impulsu elektrycznego (synapsy elektryczne). Synapsy otoczone są przez astrocyty, które uczestniczą w przekazywaniu sygnału między neuronami. Powtarzające się przekazywanie impulsu długotrwale wzmacnia przewodnictwo synaptyczne. Powstawanie sieci neuronów o zwiększonym przewodnictwie jest podstawą uczenia się i zapamiętywania. Nowe synapsy powstają w procesie zwanym synaptogenezą.

TYPY SYNAPS:

 |
| Źródło: Wikipedia. Schemat budowy koneksonu i koneksyny. |
 |
| Źródło: Wikipedia. Porównanie synapsy elektrycznej (A) i chemicznej (B); A) pobudzenie (1) jest przekazywane między komórkami za pomocą transportu jonów (2) przez bezpośrednie kanały; B) pobudzenie (1) skutkuje wydzielaniem pęcherzyków i egzocytotycznym wydzielaniem (2) substancji przekaźnikowej (3) wchłanianej endocytotycznie (4) przez komórkę - odbiornik. |

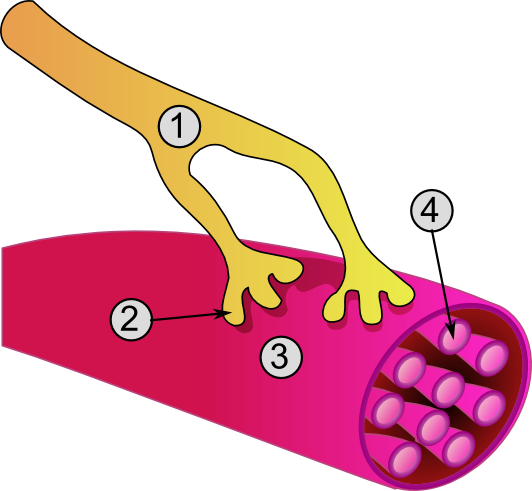 |
| Źródło: Wikipedia. Połączenie nerwowo-mięśniowe: 1 - akson, 2 - połączenie synaptyczne, 3 - mięsień szkieletowy, 4 - miofibryla. |

 |
| Źródło: Wikipedia. Struktura typowej synapsy chemicznej. |
- SYNAPTOGENEZA -> jest to proces formowania się synaps, czyli połączeń między komórkami nerwowymi. Synapsa powstaje w wyniku dwukierunkowych oddziaływań pomiędzy zakończeniem aksonu komórki nerwowej, a błoną komórki docelowej (innej komórki nerwowej lub komórki efektorowej, np. komórki mięśniowej), do której zakończenie to dociera w procesie zwanym wzrastaniem aksonów. Neurony, którym nie uda się wytworzyć połączeń, obumierają w procesie apoptozy, ponieważ nie otrzymują niezbędnych im do przeżycia czynników troficznych wytwarzanych przez komórki docelowe. Największe nasilenie synaptogenezy obserwuje się we wczesnym okresie rozwoju (przed narodzeniem) podczas kształtowania się układu nerwowego, ale tworzenie się nowych synaps w układzie nerwowym zachodzi przez całe życie osobnika - proces ten leży u podstawy zjawiska zwanego neuroplastycznością.
- POWSTAWANIE SYNAPS NERWOWO-MIĘŚNIOWYCH: Pierwszym etapem formowania się synapsy nerwowo-mięśniowej wydzielanie białka zwanego agryną przez motoneuron [u kręgowców motoneurony, zwane także neuronami motorycznymi lub ruchowymi to neurony eferentne, które wychodzą swoimi wypustkami z rdzenia kręgowego lub pnia mózgu i tworzą synapsy z włóknami mięśniowymi (motoneurony alfa) służące do wywoływania skurczu, a także synapsy z wrzecionami mięsniowymi (motoneurony gamma) słuzące do regulacji czułości proprioceptywnej] docierający do błony komórki mięśniowej. Cząsteczki agryny wiążą się z receptorami w błonie komórki mięśniowej mającymi aktywność kinaz - są to tzw. kinazy specyficzne dla mięśni (ang. muscle specific kinase, MuSK). Po związaniu cząsteczek agryny oddziaływują one z białkiem błonowym zwanym rapsyną. Aktywowane cząsteczki rapsyny indukują powstawanie skupisk receptorów nikotynowych acetylocholiny w błonie komórki mięśniowej naprzeciwko motoneuronu. Zakończenia motoneuronu wyzielają również białko zwane neuroreguliną, które wiąże się z odpowiednimi receptorami w błonie komórki mięśniowej i wywołuje kaskadę sygnałową prowdzącą do zwiększenia poziomu transkrypcji genów kodujących receptorów nikotynowych acetylocholiny i tym samym wzrost liczby tych receptorów w błonie. Wydzielana przez zakońćzenia motoneuronu acetylocholina wiąże się ze swoimi receptorami w błonie komórki mięśniowej i wywołuje jej depolaryzację. Prowadzi to m.in. do zmniejszenia transkrypcji genów kodujących receptory acetylocholiny w jądrach komórkowych położonych daleko od obszaru synaptycznego. Dzięki temu receptory leżą przede wszystkim w skupiskach w obrębie synapsy nerwowo-mięśniowej. W przypadku mięśni odnerwionych przez długi czas brak przekaźnictwa acetylocholiny wywołuje równomierne rozmieszczenie receptorów dla acetylocholiny, co jest podłożem tzw. nadwrażliwości po odnerwieniu [Prawo Cannona-Rosenblutha - jest to opisana przez Waltera Cannona i Arturo Rosenbluetha prawidłowość, głosząca, że po odnerwieniu wzrasta wrażliwość tkanek na neuroprzekaźniki]. We wczesnym okresie rozwoju jedna komórka mięśniowa może być unerwiana przez wiele motoneuronów. Ostatecznie jednak wszystkie synapsy poza tymi pochodzącymi z rozgałęzień jednego motoneuronu są eliminowane, tak że u dorosłych osobników określony motoneuron ma wyłączną kontrolę nad określoną grupą włókien mięśniowych (jest to tzw. jednostka motoryczna). Proces eliminacji nadmiaru synaps zaczyna się od zmniejszenia liczby receptorów znajdujących się w obrębie synapsy, po czym następuje wycofywanie zakończeń motoneuronu. Eliminacja lub pozostawienie danej synapsy jest wynikiem konkurencji pomiędzy zakończeniami poszczególnych motoneuronów, ale molekularne podłoże tego zjawiska nie jest jeszcze znane.
 |
| Źródło: Wikipedia. |
POWSTAWANIE SYNAPS NERWOWO-NERWOWYCH: Proces formowania się synaps nerwowo-nerwowych w środkowym układzie nerwowym jest poznany w dużo mniejszym stopniu. Uczestniczą w nim inne cząsteczki sygnałowe, w synapsach tego typu nie stwierdzono np. obecności receptorów MuSK oraz rapsyny. W procesie tym ważną rolę odgrywają znajdujące się na dendrytach wypustki zwane filopodiami (będące właściwie jedną z form morfologicznych kolców dendrytycznych), które ulegają naprzemiennemu wydłużaniu się i cofaniu aż do momentu, kiedy napotkają akson, z którym mogą wytworzyć połączenie. Dwukierunkowe oddziaływania pomiędzy błoną presynaptyczną aksonu i błoną postsynaptyczną filopodium prowadzi do tworzenia się skupisk odpowiednich receptorów (np. receptorów NMDA dla glutaminianu) w obrębie synapsy oraz wydzielania białek adhezyjnych zapewniających ścisłe przyleganie do siebie obu komponentów synapsy.
NEUROGENEZA (ang. neurogenesis) - jest to proces powstawania nowych komórek nerwowych obejmujący zarówno neurogenezę wieku embrionalnego (prenatalnego) jak i neurogenezę wieku dorosłego (postnatalnego). Neurony - główny element tkanki nerwowej swój rodowód wywodzą z komórek płytki nerwowej formowanej z ektodermy we wczesnych fazach rozwoju organizmu. Neurogeneza u ssaków ograniczona jest głównie do okresu prenatalnego i w większości przypadków zachodzi w obrębie przestrzeni okołokomorowej w której powstają neurony formujące korę nową, móżdżek i inne struktury układu nerwowego (Jacobson i in. 1991). Niemniej jednak wiele badań dowodzi, że również w dorosłym móżgu dochodzi do neurogenezy, choć na zdecydowanie niższym poziomie niż w okresie rozwoju zarodkowego (Eriksson PS 1998, Gage FK 1998).
 |
| Źródło: neurogenesis101.weebly.com |


FUNKCJONOWANIE NEURONÓW: Podstawową funkcją neuronów jest przenoszenie i przetwarzanie informacji w postaci impulsów nerwowych (potencjałów czynnościowych), będących krótkotrwałymi, gwałtownymi zmianami potencjału błony komórkowej neuronu. Impulsy nerwowe w warunkach naturalnych są przewodzone tylko w jednym kierunku [ortodromowo; przewodzenie ortodromowe to sposób przekazywania bodźca w komórce nerwowej polegający na przekazie od ciała komórki (perykarionu) do synaps. Przeciwieństwem jest drugi sposób - przewodzenie antydromowe polegające na transporcie z synaps do ciała komórki - perykarionu] od początkowego segmentu aksonu do synaps znajdujących się na jego zakończeniach. Funkcjonalnie neuron można podzielić na cztery strefy:
- strefa wejścia – dendryty i ciało komórki, które odbierają impulsy od innych neuronów poprzez znajdujące się na nich synapsy
- strefa inicjacji – początkowy odcinek aksonu, tutaj powstaje potencjał czynnościowy neuronu
- strefa przewodzenia – akson
- strefa wyjścia – synapsy na zakończeniach aksonu
 |
| Źródło: Wikipedia. Potencjał czynnościowy jest krótką i gwałtowną zmianą potencjału (odwróceniem polaryzacji elektrycznej) błony komórkowej neuronu. Powstaje w początkowym odcinku aksonu i jest przewodzony do synaps znajdujących się na zakończeniach aksonu. |
| Źródło: Wikipedia. Schemat przedstawiający zmiany wartości potencjału błonowego komórki nerwowej podczas trwania potencjału czynnościowego.
POTENCJAŁ SPOCZYNKOWY: Jest to różnica potencjałów (napięcie) między obiema stronami błony plazmatycznej niepobudzonej komórki pobudliwej. Wszystkie napięcia na błonie wyraża się jako różnice potencjału wnętrza komórki do potencjału po stronie zewnętrznej. Potencjałī spoczynkowe mają wartości ujemne, które w komórkach nerwowych wahają się między -65 mV a -90 mV. Powstanie potencjału przepływania zgodnie z gradientem stężenia tych jonów z wnętrza na zewnątrz błony komórkowej. Powoduje to pozostanie niewielkiego nadmiaru ładunków ujemnych po wewnętrznej stronie błony. Inne jony (np. sodu) jedynie w niewielkim stopniu wpływają na wartości potencjału spoczynkowego. Siła elektrochemiczna, powodująca ruch jony poprzez błonę komórkową, jest różnicą między potencjałem spoczynkowym a potencjałem równowagi dla danego jonu. Potencjał równowagi jonu jest to taki potencjał, przy którym wypływ tego rodzaju jonów z komórki jest równy ich wpływowi do jej wnętrza. Potencjał równowagi dla poszczególnych jonów można obliczyć stosując równanie Nernsta. Potencjały spoczynkowe można obliczyć stosując równanie Goldmana, które uwzględnia wszystkie zaangażowane rodzaje jonów.
  |
 |
| Źródło: Wikipedia. A -Wyidealizowany obraz potencjału czynnościowego (iglicowego), pokazujący jego poszczególne fazy; B - rejestrowane różnymi technikami rzeczywiste potencjały czynnościowe mogą odbiegać od idealnego wzorca. |
Miejscem powstawania potencjału czynnościowego w komórce nerwowej jest tzw. wzgórek aksonowy, skąd potencjał iglicowy rozprzestrzenia się po powierzchni błony komórkowej wypustki osiowej - aksonu. Generowaniem potencjałów czynnościowych rządzi zasada: "wszystko albo nic": do zapoczątkowania potencjału czynościowego potrzebny jest bodziec o intensywności wystarczającej do zdepolaryzowania neuronu powyżej określonej wartości progowej; wszystkie potencjały czynnościowe w danej komórce osiągają tę samą amplitudę.
PRZEBIEG POTENCJAŁU CZYNNOŚCIOWEGO W KOMÓRCE NERWOWEJ: Jeżeli dokomórkowe prądy kationów przeważają nad odkomórkowymi, następuje depolaryzacja od poziomu potencjału spoczynkowego (ok. -70 mV) do potencjału krytycznego czyli progowego (ok. -50 mV). Po osiągnięciu potencjału progowego, następuje otwarcie bramkowanych elektrycznie (zależne od napięcia) kanałów przewodzących odkomórkowo kationy potasowe oraz kanałó przewodzących kationy sodowe do wnętrza komórki. Wskutek różnicy stężeń i potencjałów kationy sodowe szybko napływają do wnętrza komórki, niwelując różnicę potencjałów pomiędzy środowiskiem zewnątrz- i wewnątrzkomórkowym do zera (depolaryzują błonę komórkową). Jest to początek tzw. potencjału iglicowego. Ze względu na dużą początkową różnicę stężeń, mimo wyrównania potencjałów, kationy sodowe napływają w dalszym ciągu do wnętrza komórki, powodując odwrotną polaryzację błony, tzw. nadstrzał dochodzący do +35 mV. Następuje to stosunkowo szybko; w ciągu ok 2 ms następuje już całkowita inaktywacja kanałów sodowych. Po rozpoczęciu aktywacji sodowej, w odpowiedzi na depolaryzację, następuje aktywacja potasowa, czyli otwarcie kanałów dla odkomórkowego prądu kationów potasowych. Powoduje to zmianę potencjału wnętrza komórki z powrotem na ujemny, czyli repolaryzację. W tym czasie nie jest możliwe ponowne pobudzenie komórki, jest to okres refrakcji bewzględnej [reflaksja to właściwość komórek pobudliwych, a także błon komórkowych wypustek tych komórek, polegająca na okresowej niewrażliwości na stymulujące je bodźce po przejściu potencjału czynnościowego: refkracja bezwzględna to stan, w którym komórka nie jest w stanie odpowiedzieć na żaden bodziec - tak jest zaraz po przejściu potencjału, a z czasem przechodzi w --> refrakcję względną - gdy bodziec o większym niż fizjologicznie nasileniu może wywołać odpowiedź komórki]. W czasie trwania potencjału iglicowego, a zwłaszcza pod koniec - proporcje kationów po obu stronach błony są odwrócone (Na+ wewnątrz, K+ na zewnątrz komórki). Przez cały czas trwania potencjału czynnościowego, aktywna jest pompa sodowo-potasowa oraz inne układy transportujące jony. Poziom aktywności pompy sodowo-potasowej jest bowiem uzależniony m.in. od stężenia kationów sodowych we wnętrzu komórki. Zatem w czasie trwania potencjału iglicowego pompa działa z najwyższą możliwą prędkością. W ciągu kolejnych kilkudziesięciu milisekund przywraca proporcje kationów sodowych i potasowych po obu stronach błony komókowej. Jednak początkowo błona jest jeszcze w stanie hiperpolaryzacji - różnica potencjałów przekracza wartość spoczynkową. Jest to okres refrakcji względnej, kiedy komórkę da się pobudzić, ale dużo większym bodźcem - ze względu na większą "odległość" od poziomu potencjału krytycznego. W pewnym stopniu, w różnych tkankach, w opisanych procesach biorą udział również inne jony, głównie jony chlorkowe (Cl-) oraz wapniowe (Ca2+).
ZASADA WSZYSTKO ALBO NIC: Przewodzenie sygnałów przez neuron podlega zasadzie wszystko albo nic, tzn. neuron wytwarza potencjał czynnościowy lub nie. Wszystkie powstające potencjały czynnościowe w danej komórce nerwowej mają tę samą wielkość, bez względu na wielkość bodźca (o ile tylko jest on wystarczająco intensywny, aby wywołać powstanie potencjału). Silniejsza stymulacja neuronów (bodźcami ponadprogowymi) nie prowadzi do wytwarzania silniejszych potencjałów. Może natomiast prowadzić do zwiększenia częstotliwości wytwarzania potencjałów przez neuron.
 |
| Źródło: wellcomeimages.org - John Zachary Young. Wellcome Institute. |

- The Squid and its giant nerve fibre - Part 1
- The Squid and its giant nerve fibre - Part 2
 |
| Źródło: neuronbank.org - Gigantyczny akson kalmara. |
- "The Squid and its Giant Nerve Fiber" - seria filmów, Plymouth Marine Laboratory - KLIK!
 |
| Źródło: nanoHUB.org - Osiągnięcia naukowe Hodgkina i Huxleya.
SYGNAŁ WIDOCZNY NA OSCYLOSKOPIE: Pokazuje to, co Alan Lloyd Hodgkin i Andrew Huxley zaobserwowali w latach 40. XX wieku w gigantycznym aksonie kałamarnicy. W 1952 roku opisali swój model mechanizmów jonowych leżących u podstaw inicjacji i propagacji potencjałów czynnościowych. Został nazwany modelem Hodgkina-Huxleya i za tę pracę otrzymał Nagrodę Nobla z 1963 roku w dziedzinie fizjologii lub medycyny. Sygnał jest tutaj z jednego pojedynczego C-włókna ssaka.
MODEL PROPAGACJI IMPULSU: Wówczas, gdy dendryt odbierze jeden z bodźców z drugiej strony synapsy oddzielającej akson od dendrytu, przez błonę dendrytu zaczynają przepływać miniaturowe prądy elektryczne. Zazwyczaj są to prądy wpływające do wnętrza dendrytu, określane jako pobudzające. Prądy mogą także wypływać na zewnątrz komórki i wówczas noszą nazwę prądów hamujących. Zarówno dodanie, jak i ujemne wahania prądu, powstające w błonach dendrytów, przepływają do ciała komórkowego. Jeżeli ich suma jest niewielka, prądy takie zanikają i nie uruchamiają dalszych procesów. Jednak jeśli w wyniku ich dodawania zostanie przekroczona określona wartość progowa, to generowany jest tzw. potencjał czynnościowy propagujący się wzdłuż aksonu i komórka nerwowa wysyła wiadomość do innych neuronów. To działa prawie tak jak komputer: jest impuls lub impulsu brak. Jeżeli jest impuls to jest na plus (czyli +1), jeżeli nie ma impulsu to jest na minus (czyli -1). Logika 2-wartościowa (+1, -1) potrafi zrobić wszystko: potrafi zakodować wszelkie informacje, potrafi wykonywać skomplikowane operacje, a nawet potrafi się uczyć (przykłady rozmaitych algorytmów). To właśnie na tym polega fenomen komórek nerwowych. Model wyjaśniający działanie neuronów opisali angielscy uczeni - Hodgkin i Huxley w 1952 roku. Jest to model opisujący działanie neuronu w sposób jakościowy.
Neuron i jego otoczenie wypełnione jest płynem zawierającym jony: potasu (K+), sodu (Na+), wapnia (Ca2+) i chloru (Cl-). Pompy jonowe wypompowują z komórki jony sodu, a wpompowują jony potasu. Na zewnątrz komórki istnieje wysoka koncentracja Na+, Cl- i Ca2+. Wewnątrz istnieje wysokie stężenie K+ oraz ujemnie naładowanych molekuł. Membrana komórkowa, oprócz pomp jonowych, zawiera kanały jonowe. W stanie równowagi, zewnętrzna część błony komórkowej naładowana jest dodatnio, a wewnętrzna ujemnie. Różnicę potencjałów nazywamy potencjałem spoczynkowym. Typowo waha się on pomiędzy -90 mV a -65 mV. Jeżeli sygnał pobudzający neuron przekroczy wartość progową, w neuronie nastąpi generacja tzw. potencjału czynnościowego o wartości około +40 mV i sygnał rozchodzi się w aksonie w postaci fali depolaryzacji i następującej po niej repolaryzacji o stałej czasowej 1 ms.
Przez V oznaczamy różnicę potencjałów pomiędzy wewnętrzną a zewnętrzną stroną membrany neuronu. Gradient stężeń danego typu jonów "k" powoduje powstanie różnicy potencjałów Ek (nazywanego potencjałem Nernsta). Wypadkowym potencjałem jest: V−Ek ---> potencjał elektrochemiczny. Z praw Ohma otrzymujemy relację dla natężenia prądu danego typu jonów:
Ik=V−EkRk=Gk(V−Ek)
    
Źródło: visual.icse.us.edu.pl
Hodgkin-Huxley MODEL - Wikipedia (EN) - KLIK!
|
 |
| Źródło: Wikimedia. Budowa typowej komórki nerwowej.
Dendryty - osiągają one długość do 1 mm. Rozgałęziają się na całej swej długości, a w miarę oddalania od somy zmniejsza się ich średnica. Są otoczone błoną komórkową składającą się z mikrotubuli, neurofilamentów i cytosolu, pod którą znajduje się cytoplazma. Ze względu na to, że dendryty stanowią przedłużenie cieła komórki, można w nich znaleźć organelle tam występujące - mitochondria, szorstkie retikulum endoplazmatyczne, aparat Golgiego, rybosomy. W niektórych neuronach dendryty są pokryte cienkimi tworami zwanymi kolcami dendrytycznymi, na których szczycie tworzą się synapsy z innymi neuronami. Komórki nerwowe zawierające kolce nazywane są neuronami kolczastymi, a te, które ich nie mają - neuronami bezkolcowymi. Rozgałęzienia dendrytowe mogą rozrastać się lub kurczyć. Alkohol wpływa szkodliwie na wzrost dendrytów, w podeszłym wieku zaś dochodzi do skrócenia i zmniejszenia liczby tych rozgałęzień.
Funkcje: dendryty są wyspecjalizowane w odbieraniu bodźców i przesyłaniu sygnałów do ciała komórkowego, które integruje dochodzące sygnały. Impulsy nerwowe są przewodzone wzdłuż błony komórkowej. Wytwarzając połączenia z innymi neuronami i przewodząc impulsy, dendryty pełnią funkcje integrujące czynności wielu komórek nerwowych. Pojedyncze, długie dendryty nerwów rdzeniowych przewodzą impulsy elektryczne wzbudzone bodźcami czuciowymi do ciał komórkowych neuronów czuciowych. Dendryty znacznie zwiększają powierzchnię komórek nerwowych - stanowią do 90% powierzchni wielu neuronów.
MORFOLOGIA: Obserwuje się różnice w rozmiarach osiąganych przez kolce dendrytyczne, zarówno między różnymi gatunkami jak i w różnych obszarach mózgu. Kolce w kroze mózgu człowieka osiągają zwykle długość ok. 1 mikrometra. Typowy, dojrzały kolec składa się z cienkiej, odchodzącej od dendrytu "szyjki" o średnicy ok. 0,1 mikrometra oraz szerszej "główki" o średnicy ok. 0,5 mikrometra. Wyróżnia się trzy podstawowe typy budowy kolca:
Pomiędzy tymi podstawowymi typami budowy obserwuje się formy pośrednie. Ponadto na rozwijających się dendrytach występują bardzo cienkie i długie, pozbawione główki filopodia będące formą niedojrzałą kolców. Niektóre kolce tworzą rozgałęzienia (ang. branched spines) i mają więcej niż jedną synapsę na szczycie.

Źródło: synapses.clm.utexas.edu - Trójwymiarowa rekonstrukcja kolców dendrytycznych. Kolorem czerwonym zaznaczono synapsy utworzone na kolcach; kolorem niebieskim - synapsy znajdujące się bezpośrednio na dendrycie.
BUDOWA WEWNĘTRZNA: Za dużą plastyczność kolców odpowiadają filamenty aktynowe będące głównym składnikiem ich cytoszkieletu. W kolcach dendrytycznych obecne są liczne rybosomy co świadczy o wysokim poziomie syntezy białka zachodzącej w tych strukturach. Nie występują natomiast mikrotubule oraz mitochondria. Około 10% powierzchni błony komórkowej kolców wchodzi w skład synaps i zawiera receptory dla glutaminianu (głównego neuroprzekaźnika pobudzającego) - są to m.in. receptory NMDA oraz receptory AMPA. Ponadto w błonie komórkowej kolców obecne są również receptory dla BDNF - czynnika wzrostu neuronów.

Źródło: Dr, JohnC. Fiala, and Dr. Kristen M. Harris - Atlas of Ultrastructural Neurocytology - Trójwymiarowa rekonstrukcja pojedynczego kolca dendrytycznego. Kolorem czerwonym oznaczono synapsy, a kolorem czarnym rybosomy.
ROZWÓJ: Kolce dendrytyczne rozwijają się na dendrytach ssaków w pierwszych tygodniach życia. Najszybszy wzrost liczby kolców obserwuje się w drugim i trzecim tygodniu życia. Początkowo na dendrytach powstają filipodia - cienkie, pozbawione główki wypustki, które nie tworzą synapsy. Stopniowo przekształcają się one w dojrzałe kolce. U młodych osobników okres "półtrwania" kolca jest dość krótki - tempo powstawania i eliminacji kolców dendrytycznych zachodzi na stosunkowo wysokim poziomie. U dorosłych osobników okres ten się skraca i obserwuje się większą stabilność kolców. Ponadto podczas starzenia się zachodzi stopniowy spadek ogólnej liczby kolców. Różne zaburzenia rozwoju skutkujące wystąpieniem, upośledzenia umysłowego (np. zespół Downa, alkoholowy zespół płodowy) wiążą się z nieprawidłowym rozwojem kolców dendrytycznych.
Akson (Neuryt, włókno nerwowe) - jest to element neuronu odpowiedzialny za przekazywanie informacji z ciała komórki do kolejnych neuronów lub komórek efektorowych (np. komórek mięśniowych bądź gruczołowych). Neuryt może być osłonięty osłonką włókien nerwowych. Jest z reguły dłuższy od dendrytów i, w odróżnieniu od nich, w komórce występuje pojedynczo (chociaż może być on rozgałęziony). Czasem wyróżnia się włókna nerwowe od neurytów nazywają włóknami nerwowymi długie neuryty neuronów. Czasami włóknami nerwowymi określa się także dowolne wypustki neuronu.

*Włókno czuciowe, włókno nerwowe aferentne, włókno nerwowe dośrodkowe - jest to włókno nerwowe przewodzące impulsy nerwowe do ośrodkowego układu nerwowego od receptoró unerwiających narządy wewnętrzne, mięśnie szkieletowe, skórę i stawy.

Źródło: docplayer.pl - "Tkanka nerwowa, centralny układ nerwowy" - prezentacja użytkownika Bartłomiej Mróz.
Rozchodzenie się impulsu nerwowego wzdłuż aksonu: Generowanie i rozchodzenie się impulsu nerwowego związane są z transportem jonów. Przy ciągłym przepływie jonów przez błonę konieczny jest nakład energii.
Otoczka mielinowa - jest to osłonka włókien nerwowych tworzona przez oligodendrocyty w ośrodkowym układzie nerwowym i przez komórki Schwanna w obwodowym układzie nerwowym. Komórki, owijając się kilkakrotnie wokół neurytów, tworzą osłonkę z własnej błony komórkowej, ułożonej w kilka warstwa złączonych białkiem PLP1. Spełnia ona jednocześnie funkcję ochrony mechanicznej i izolatora elektrycznego aksonu. Długie aksony z osłonką mielinową nazywane są włóknami rdzennymi, a aksony bez osłonki to włókna bezrdzenne. Osłonkę mielinową ma większość długich neurytów, biegnących przez substancję białą w ośrodkowym układzie nerwowym oraz w nerwach rdzeniowych. W odstępach około 1 mm w obrębie cieśni węzła (przewężeń Ranviera) włókna rdzenne pozbawione są osłonek.
Mielina to substancja, która wytwarzana jest przez komórki otaczające aksony komórek nerwowych. W ośrodkowym układzie nerwowym są to oligodendrocyty, natomiast w obwodowym układzie nerwowym są to komórki Schwanna (lemocyty). Głównym składnikiem osłonki mielinowej są cerebrozydy, a w tym galaktozyloceramid. Związki te są zaliczane do grupy glikolipidów; składają się z cukrów; w tym przypadku galaktozy i lipidu - ceramidu (pochodna sfingozyny). Innym składnikiem mieliny jest fosfolipid lecytyna (fosfatydylocholina) + inne składniki: zasadowe białko mieliny (MBP - 30% białek mieliny), mielinowa glikoproteina oligodendrocytów, proteina proteolipidowa (PLP, ok. 50% białka mieliny); CHOLESTEROL.
*Splatanie łańcuchów węglowodorowych sfingomieliny służy do wzmocnienia mieliny.

Źródło: Wikipedia.

Źródło: edunauka.pl - Cerebrozyd - schemat budowy cząsteczki.
 |


Źródło: stwardnienieforum.pl
 |
| Źródło: Wikipedia. Przewężenia Ranviera - wybarwione na zarno. |
Przewężenia Ranviera - są to przerwy, które występują między komórkami osłonki mielinowej. W przewężeniu zostaje wzmocniony impuls sygnału nerwowego. Osłonka mielinowa wymusza przepływ impulsów od przewężenia do przewężenia pełniąc rolę elektrycznej izolacji przyspieszającej sygnał elektryczny. W tych przewężeniach błona komórkowa aksonu (zawierająca gęsto rozmieszczone kanały jonowe) nie jest osłonięta mieliną, a więc kontaktuje się z płynem pozakomórkowym. Dzięki temu prąd przenosi się skokowo od węzła zdepolaryzowanego do węzła będącego w spoczynku - impulsy nerwowe skaczą. Przewodzenie jest skokowe (saltacyjne, łac. saltare) jest znacznie szybsze niż przewodzenie ciągłe we włóknach niezmielinizowanych. Potencjał czynnościowy sięga do 140 mV. Przewężenia mają długość rzędu 1 mikrometra i poprzedzielane są mniej więcej regularnie odcinkiem zmielinizowanym o długości 50 mikrometrów - 1 mm. Gradient potencjału w przewężeniu ejst znacznie większy niż w zmielinizowanym odcinku aksonu.
O Louisie-Antoine Ranvierze możesz przeczytać: tutaj.
RÓŻNORODNOŚĆ I KLASYFIKACJA NEURONÓW:
Typy neuronów -> KLIK!
RÓŻNE TYPY NEURONÓW MOŻESZ OBEJRZEĆ NA TEJ STRONIE (KLIK!).
W mózgu człowieka jest ponad 10000 specyficznych typów neuronów, ale ogólnie wyróżniamy trzy główne typy neuronów ze względu na pełnione na nie funkcje: neurony ruchowe (przekazują informacje motoryczne), neurony czuciowe (przekazują informacje czuciowe) i interneurony (przekazują informacje różnych typów komórek nerwowych). Neurony mogą także przyjmować różne kształty i rozmiary (zobacz to na poniższych rysunkach wykonanych przez Cajala - hiszpańskiego histologa, neuroanatoma, prekursora neurobiologii, który wraz z Camillo Golgim nagrodzony został Nagrodą Nobla w dziedzinie medycyny w roku 1906).


 |
| Źródło: Wikipedia. Neuron piramidowy z hipokampu człowieka (barwiony metodą Golgiego). |
- --> Neurony piramidowe - jest to morfologiczny typ neuronu, którego cechą charakterystyczną jest ciało komórki o kształcie trójkąta oraz obecność dwóch odrębnych drzewek dendrytycznych. Neurony piramidowe leżące w korze ruchowej mózgu są częścią układu piramidowego i biorą udział w kontroli ruchów dowolnych i postawy ciała. Ponadto neurony piramidowe leżące w korze przedczołowej są zaangażowane w szereg procesów poznawczych, m.in. w funkcjonowanie pamięci roboczej. WYSTĘPOWANIE -> obecność neuronów piramidowych w mózgu zaobserwowano u wszystkich badanych dotąd ssaków i u innych kręgowców (ryb, ptaków, gadów) z wyjątkiem płazów. Neurony piramidowe stanowią ok. 60% wszystkich neuronów kory mózgowej. W II, III oraz VI warstwie kory znajdują się małe neurony piramidowe, a w V warstwie duże komórki piramidowe zwane komórkami Betza. Poza korą mózgową neurony piramidowe są obecne również w hipokampie (w warstwie CA1 i CA3) oraz w ciele migdałowatym. STRUKTURA -> cechą charakterystyczną neuronów piramidowych (od której powstała ich nazwa) jest trójkątny kształt ich ciała komórki. Od innych neuronów odróżnia ich również obecność dwóch odrębnych drzewek dendrytycznych: apikalnego i bazalnego. Drzewko apikalne składa się z długiego dendrytu apikalnego, który odchodzi od wierzchołka ciała komórki i w pewnej odległości od niej tworzy szereg rozgałęzień. Drzewko bazalne składa się z dendrytów odchodzących bezpośrednio od podstawy ciała komórki, które również wielokrotnie rozgałęziają się. Dendryty bazalne są dużo krótsze od dendrytu apikalnego, mają również dużo mniejszą średnicę. Od podstawy ciała komórki odchodzi również pojedynczy akson. Wszystkie dendryty neuronów piramidowych są pokryte licznymi kolcami dendrytycznymi. FUNKCJE -> duże neurony piramidowe leżące w V warstwie pierwszorzędowej kory ruchowej (zwane komórkami Betza) biorą udział w kontroli ruchów dowolnych i postawy ciała - stanowią początek układu piramidowego. Ich aksony przechodzą przez torebkę wewnętrzną i biegną do pnia mózgu, gdzie część z nich tworzy połączenia synaptyczne z motoneuronami unerwiającymi m.in. mięśnie twarzy. Pozostałe aksony biegną dalej przez rdzeń przedłużony i wzdłuż rdzenia kręgowego, gdzie tworzą połączenia synaptyczne z motoneuronami unerwiającymi pozostałe mięśnie (m.in. mięśnie kończyn). Neurony piramidowe leżące w korze przedczołowej są zaangażowane w procesy poznawcze, m.in. w funkcjonowanie pamięci roboczej.
 |
| Źródło: Wikipedia. Neuron piramidowy z kory mózgowej myszy - uwidoczniony dzięki zastosowaniu białka GFP - białko zielonej fluorescencji. |
- Komórki Betza (Olbrzymie komórki piramidowe) - jest to rodzaj neuronów piramidowych zlokalizowanych w V warstwie istoty szarej pierwszorzędowej kory ruchowej (M1). Zostały one opisane w 1874 roku przez ukraińskiego anatoma i histologa Wladimira Betza. Są to jedne z największych komórek ośrodkowego układu nerwowego. Ich perikariony mają nawet 100 mikrometrów średnicy. Komórki Betza wysyłają aksony do rdzenia kręgowego, gdzie tworzą synapsy z komórkami rogów przednich, stanowią zatem pierwszy neuron dróg piramidowych (nazwa komórek piramidowych wzięła się jednak od ich kształtu, zanim poznano dokładny przebieg dróg piramidowych). Komórki Betza mają jeden apikalny dendryt, jak inne komórki piramidowe, poza nim mają też licznie odgałęziające się w różnych komórkach asymetryczne dendryty z których większość dochodzi do warstw V i VI kory, a także głębiej, do istoty białej mózgu.
 |
| Źródło: D.L. Felten - "Atlas neuroanatomii i neurofizjologii Netlera" 2003 - Komórka Betza. |
--> Komórki Purkiniego (opis zamieszczony w dalszej części omówienia)
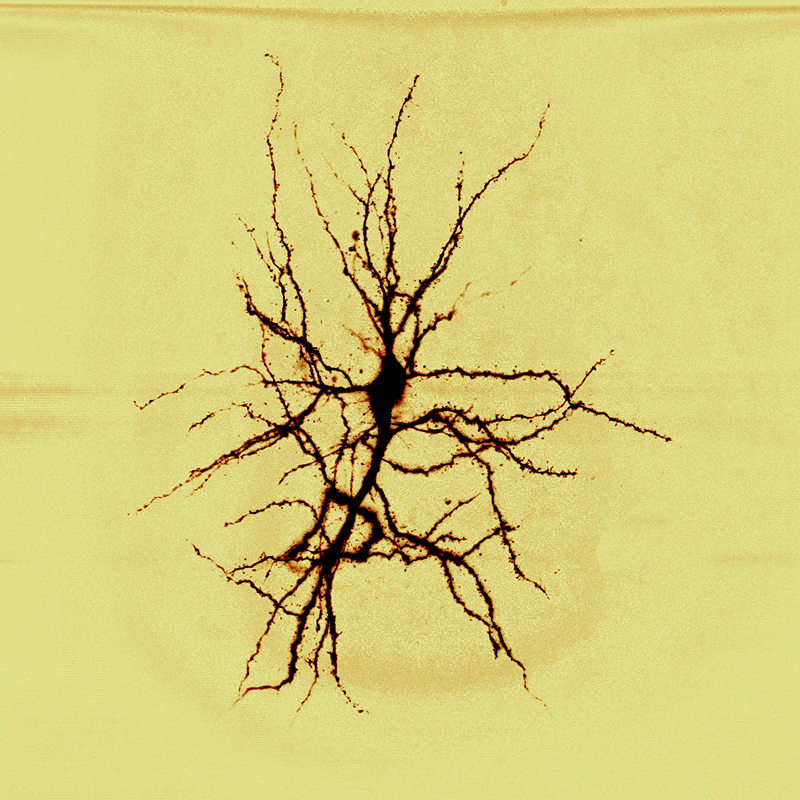 |
| Źródło: brain.utah.edu - Komórka gwiaździsta z kory śródwęchowej. Uwidoczniona została dzięki zastosowaniu barwnika fluorescencyjnego Alexa 488. |
--> Komórki gwiaździste, neurony gwiaździste (ang. stellate cell) - jest to morfologiczny typ neurony, który charakteryzuje się radialnym ułożeniem dendrytów nadającym mu kształt gwiazdy. Komórki gwiaździste pełnią rolę interneuronów czyli przekazują impulsy nerwowe pomiędzy neuronami, na niewielką odległość. Występują m.in. w warstwie drobinowej kory móżdżku gdzie tworzą synapsy na dendrytach komórek Purkiniego i działają na nie hamująco przez uwalnianie neuroprzekaźnika GABA; a także w warstwie 4C w pierwszorzędowej korze wzrokowej, gdzie pośredniczą w przekazywaniu informacji wzrokowej od wzgórza do wyższych warstw kory: otrzymują sygnały pobudzające od komórek zwojowych P i M ciała kolankowatego bocznego wzgórza i same również działają pobudzająco na neurony piramidowe warstwy 4B.
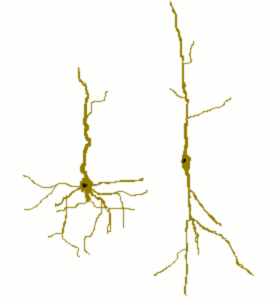 |
| Źródło: Wikipedia. Schematyczne przedstawienie zwykłego neuronu (po lewej) i neuronu von Economo (po prawej). |
--> Neurony von Economo / Neurony wrzecionowate (ang. spindle neurons, von Economo neurons, VENs) - jest to rodzaj komórek nerwowych, charakteryzujący się wrzecionowatym perikarionem, z jednej strony przechodzącym w pojedynczy neuryt, po drugiej stronie w pojedynczy dendryt. Neurony von Economo znajdują się w określonych obszarach kory mózgowej ludzi i innych człowiekowatych, waleni (humbaków, finwali, orek, kaszalotów, butlonosów, delfinów Risso, białuch), a także słoni afrykańskich i indyjskich. Odkrywcą tych szczególnych neuronów był Constantin von Economo. U ludzi i małp neurony von Economo znajdują się w korze przedniego zakrętu obręczy (ACC) i czołowej kory wyspowej (FI). Niedawno wykazano obecność neuronów o podobnej morfologii w korze przedczołowej. Ich występowanie wyłącznie u najinteligentniejszych ssaków było przesłanką do hipotezy, że są one wyrazem adaptacji neuronalnej w dużych mózgowiach, umożliwiając szybkie przetwarzanie i przekazywanie informacji, co ma związek z zachowaniami społecznymi. Zaproponowano, że dysfunkcja neuronów von Economo może mieć związek z patogenezą autyzmu. Zmniejszenie liczby tych komórek opisywano w mózgach z agenezją ciała modzelowatego i otępieniem czołowo-skroniowym.
 |
| Źródło: Wikipedia. Przekrój przez ludzki móżdżek. Widoczne duże ciała komórkowe komórek Purkiniego otoczone przez zakoń- -czenia aksonów neuronów koszyczkowych. |
--> Neurony koszyczkowe - neuron koszyczkowy to morfologiczny typ neuronu, którego zakończenia aksonu tworzą charakterystyczny kształt koszyczka wokół ciała komórki docelowego neuronu, na który działają hamująco przez wydzielanie neuroprzekaźnika GABA. Neurony koszyczkowe występują przede wszystkim w warstwie drobinowej kory móżdżku, gdzie tworzą synapsy na ciałach komórkowych komórek Purkiniego. Poza tym są również obecne w hipokampie (w warstwie CA3) oraz korze mózgu, gdzie wywierają hamujący wpływ na neurony piramidowe.
PODZIAŁ FUNKCJONALNY NEURONÓW:
--> Neurony czuciowe (aferentne, dośrodkowe) - są to neurony, które wychodzą swoimi wypustkami z włókien mięśniowych i tworzą synapsy z rdzeniem kręgowym lub pniem mózgu. W łuku odruchowym bodźce są przekazywane neuronami czuciowymi do neuronów pośredniczących, skąd odpowiedź na nie jest przesyłana neuronem motorycznym do mięśnia.
--> Neurony ruchowe, motoryczne (Motoneurony) - są to neurony eferentne, które wychodzą swoimi wypustkami z rdzenia kręgowego lub pnia mózgu i tworzą synapsy z włóknami mięśniowymi (motoneurony alfa) służące do wywoływania skurczu, a także synapsy z wrzecionami mięśniowymi (motoneurony gamma) służące do regulacji czułości proprioceptywnej. *Reinerwacja - jest to proces zachodzący w żywej tkance, w którym neurony motoryczne tworzą nowe neuryty, aby przejąć funkcję znajdujących się w pobliżu dysfunkcyjnych nerwów.
--> KOJARZENIOWE (przenoszące informajce pomiędzy neuronami, tzw. Interneurony):
 |
| Źródło: Wikimedia. Interneuron. |
 |
| Źródło: Wikipedia. Zarodek danio pręgowanego 48 godzin od zapłodnienia. Strzałki wskazują cztery neurony Rohona-Bearda. |
--> Komórki Rohona-Bearda - są to wyspecjalizowane neurony o funkcji mechanoreceptorowej występujące na etapie embrionalnym rozwoju w grzbietowej części rdzenia kręgowego u ryb i płazów. Neurony Rohona-Bearda wykształcają się na granicy ektodermy epidermalnej (powierzchniowej) i neuroektodermy, jako pierwsze w kolejności neurony receptorowe. U większości gatunków zanikają one w trakcie rozwoju osobniczego (np. u danio pręgowanego w ciągu pierwszych dwóch dni rozwoju, na etapie larwalnym, by zostać zastąpionymi przez zwój korzeni grzbietowych nerwów rdzeniowych) na drodze apoptozy. W naukach biologicznych są przedmiotem badań nad apoptozą. Opisali je niezależnie od siebie John Beard i Jozef Viktor Rohon.

LICZBA NEURONÓW: Szacuje się, że ludzki mózg zawiera ok. 1,5-1,6 x 1011 neuronów i 1014 synaps. Ogromna większość neuronów znajduje się w móżdżku, a najliczniejszą populacją komórek są małe neurony ziarniste móżdżku.
Nicień Caenorhabditis elegans, będący organizmem modelowym w biologii, posiada jedynie 302 neurony. Muszka owocowa Drosophila melanogaster posiada ok. 100 000 neuronów.
Osiągnięcia Hiszpańśkiej Szkoły Neurologicznej: Do czasu odkrycia Ramona y Cajala i Camillo Golgiego uważano, że mózg jest jednolitą masą, ponieważ metodami, jakimi dysponowano i stosowano, nie potrafiono uzyskać obrazu komórek nerwowych.
Obaj naukowcy zastosowali tę samą metodę barwienia preparatów histologicznych, która oparta była na nowej wówczas technice fotograficznej. Została ona opracowana w roku 1873 przez Golgiego (tzw. metoda Golgiego). Golgi uważał, że struktura obserwowanych tkanek ejst siecią wielu rozgałęziających, bezpośrednio połączonych nerwowych kanalików.
 |
| Źródło: Wikipedia. Metoda Golgiego. |
 |
| Źródło: robotspacebrain.com Santiago Ramon y Cajal. |
“As long as our brain is a mystery, the universe, the reflection of the structure of the brain will also be a mystery.” – Santiago Ramón y Cajal
W 2017 roku archiwum Santiago Ramona y Cayala i jego współpracowników zostało wpisane na listę Pamięci Świata jako Archives of Santiago Ramon Y Cajal and the Spanish Neurohistological School. Archiwum, zawierające prace uczonego (preparaty, rysunki, fotografie, opisy) oraz wytworzone przez niego dzieła sztuki (rysunki i obrazy), fotografie, jego korespondencję, notatki, książki (napisane oraz będące w jego bibliotece) są ważnym źródłem do poznania historii rozwoju neurobiologii.
 |
| Źródło: thebeautifulbrain.com - Ramon Cajal. |
Więcej informacji na temat Ramona Cajala znajdziesz: tutaj.
Hiszpańska szkoła neurologiczna:
 |
| Źródło: neurodrawings.com - Cajal i jego współpracownicy. Hiszpańska szkoła neurologiczna. |
Łącznie ta grupa hiszpańskich neuronaukowców skupionych przez Santiago Ramona y Cajala jest zbiorowo znana jako hiszpańska szkoła neurologiczna (lub neurologiczna), lub po prostu Szkoła Cajal i stanowi jeden z najbardziej wyjątkowych przykładów szkół naukowych w historii nauki z powodu jej licznych naukowych sukcesów i odkryć.
El Archivo de Santiago Ramón y Cajal y la Escuela Española de Neurohistología (Archives of Santiago Ramon Y Cajal and the Spanish Neurohistological School) – to zbiory książek, dokumentów i przedmiotów pozostałych po Santiago Ramonie y Cajalu - hiszpańskim pionierze neurologii oraz kilku jego uczniach i współpracownikach. W skład wchodzą książki, rękopisy, preparaty anatomiczne (głównie neurologiczne), rysunki i obrazy (zarówno ilustracje medyczne jak i dzieła artystyczne), korespondencja, zdjęcia (zarówno naukowe, jak i inne) oraz rzeczy używane przez naukowców.

 |
| Źródło: Wikipedia. Pio del Rio-Hortega w 1924 roku. |
 |
| Źródło: p.pinimg.com. Del Rio-Hortega, 1922. Structure of the pineal gland. Struktura szyszynki. |
 |
| Źródło: researchgate.net - Fernando de Castro. |
--> Archive Fernando de Castro - w posiadaniu rodziny Fernando de Castro znajdują się: 3 książki, 2 podręczniki i 65 publikacji (w tym 9 dotyczących Cajala oraz Szkoły), rękopis książki napisanej w 1933 wspólnie z Cajalem, 11 innych dokumentów i około 2 tysięcy listów. Oprócz tego 609 rysunków (w tym 531 sporządzonych przez Castro), 4129 preparatów i tysiące fotografii różnych autorów. Przechowywane są też 3 mikroskopy i inne pamiątki.
RYSUNKI FERNANDO DE CASTRO ZNAJDZIESZ: TUTAJ.
Zaraz po uzyskaniu doktoratu pracujący nad strukturą zwojów czuciowych i wegetatywnych, wraz z umiejętnościami nabytymi w tym czasie w zakresie technik histologicznych, Fernando de Castro zaczął badać unerwienie rejonu tętniczo-szyjnego. W pierwszych dwóch dekadach XX wieku rejon ten był był celem licznych badań, po pojawieniu się opisu odruchów oddechowych Heinricha Heringa. Dwa istotne badania De Castro stworzyły anatomiczne podstawy dla wyjaśnienia istoty owych odruchów (badania publikowane w 1926 i 1928 roku). W nich Fernando de Castro opisał rozdzieloną dystrybucję baroreceptorów w zatoce tętnicy szyjnej, a także chemoreceptory w ciele szyjnym (glomus caroticum). Wykazał również sensoryczną naturę tego unerwienia, oddzielonego od włókien nerwowych pochodzących z wegetatywnych zwojów. Najbardziej znacząca fizjofarmakologiczna grupa owych czasów, związana z osobą Corneille'a Heymansa (1892-1968) z Ghent University (Belgia) przedstawiła znaczenie pracy De Castro i, po zaproszeniu Fernanda de Castro do Ghent, aby wspólnie przeprowadzili różne eksperymenty, szkoła Ghent School postanowił skoncentrować się na carotid body (ciele szyjnym) w celu zbadania chemoreceptorów tętniczych. Ta decyzja była kluczowa dla ówczesnej pracy Corneille Heymansa i to on otrzymał Nagrodę Nobla z 1938 roku w dziedzinie fizjologii lub medycyny, podczas gdy Fernando de Castro znajdował się pod ciężkim zagrożeniem życia w Madrycie, broniąc Instytutu Cajal przed grabieżą podczas hiszpańskiej wojny domowej (1936-1939). Królewska Szwedzka Akademia Nauk (The Swedish Nobel Academy) zapomniała o opisie De Castro, chociaż już wielokrotnie był on przywoływany na świecie i zostął uznany za fundamentalny wkład młodego neurologa z Hiszpanii i jego zasług; pojawiały się głosy, by podzielić Nagrodę Nobla pomiędzy Heymansem i de Castro.
Fernando de Castro kontynuował swoje badania na ten temat. Ważnymi publikacjami były te opublikowane w 1940, 1944, 1950, 1951, 1962 i pośmiertelnie w 1968.

Inne przykładowe rysunki de Castro:
Źródło: neurodrawings.org


Wiele zawdzięczamy De Castro w dziedzinie badań nad komórkami glejowymi!
Rys: Ilustracja histogenezy neurogleju w opuszce węchowej (1916):
Rys: Typowe waskularne zakończenia astrocytów w opuszce węchowej (1916):
 |
| Źródło: aragonesesilustres.wikispaces.com - Pedro Ramon y Cajal. |
--> Archive Pedro Ramon y Cajal - archiwum pozostawione przez Pedro Ramona y Cajala, brata Santiago. Przechowywane przez krewnych Pedro. Składa się z 20 publikacji naukowych, 2 manuskryptów Santiago i 8 listów oraz manuskryptów Pedro, 50 rysunków oraz 150 preparatów. W skład kolekcji wchodzi też 350 płyt ze zdjęciami stereoskopowymi (fotografia stereoskopowa skłąda się z dwóch zdjęć tego samego obiektu, ale wykonanych z różnych punktów widzenia. Patrzący przez stereoskop widz ma wrażenie przestrzenności i trójwymiarowości oglądanej sceny) wykonanymi przez Santiago.
 |
| Źródło: bancodeimagenesmedicina.com - Rafael Lorente de No. |
 |
| Źródło: frontiersin.org - Nieopublikowane rysunki kory mózgowej szczura wykonane przez Lorente de No. |
PRZYKŁADOWE RYSUNKI CAJALA:
Źródła: nobelprize.org
Rys.01.
Rysunek kory móżdżku (na podstawie wyglądu preparatu móżdżku kota przygotowanego za pomocą metody barwienia Golgiego). Litera A oznacza komórki Purkiniego z charakterystycznymi dendrytycznymi rozgałęzieniami. Komórki Purkiniego znajdują się w móżdżku stanowiąc jego najważniejszy element komórkowy. Kora móżdżku jest trzywarstwowa i składa się z warstwy drobinowej, zwojowej i ziarnistej. Najbardziej powierzchowną jest warstwa drobinowa, poniżej której położona jest wąska warstwa zwojowa komórek Purkiniego. Na drzewkowato rozgałęzionych dendrytach tych komórek kończą się włókna pnące i włókna równoległe. Od podstawy komórek Purkinjego odchodzi akson biegnący przez warstwę ziarnistą do neuronów jąder móżdżku. Najgłębiej położoną warstwą kory móżdżku jest warstwa ziarnista, w której znajdują się bardzo liczne komórki ziarniste małe i rzadziej występujące duże., ziarniste komórki Golgiego. Małe komórki ziarniste charakteryzują się obecnością silnie rozgałęzionych dendrytów (na kształt szpnoów). Aksony tych komórek wędrują do komórek Purkiniego.
Komórki Purkiniego jako duże, wielobiegunowe neurony rozciągają się na wszystkie trzy warstwy. Ich silnie rozgałęzione w jednej płaszczyźnie dendryty znajdują się w warstwie drobinowej, perykariony tworzą warstwę zwojową, a akson biegnie do istoty białej móżdżku (do jądra zębatego, jądra czopowatego, kulkowatego i jądra wierzchu) poprzez warstwę ziarnistą.
 |
| Źródlo: NeuroMorpho.org - Model komórki Purkiniego. |
Dendryty apikalne komórek Purkinjego mają ogromną liczbę kolców dendrytycznych (ponad 80 000 na komórkę).
Komórki Purkinjego zawierają kwas gamma-aminomasłowy (GABA) - neurotransmiter hamujący (przez co działają hamująco na swoje miejsca docelowe). Charakterystyczną cechą tych komórek jest także ekspresja białka kalbindyny. Barwienie kalbindyny z mózgu szczura po przewlekłym jednostronnym uszkodzeniu nerwu kulszowego sugeruje, że nowe neurony Purkniego mogą być tworzone w dorosłym mózgu, inicjując organizowanie nowych płatów w móżdżku.
Rys.02.
Na rysunku tym możemy zaobserwować różnorodność form morfologicznych neuronów - widzimy np. tutaj neurony dwubiegunowe.

Rys.04.
Inne rysunki Cajala znajdziesz: tutaj.
KOMÓRKI GLEJOWE: Komórki glejowe lub glej (gr. glia = klej; ang. glial cells), stanowią obok komórek nerwowych drugi składnik tkanki nerwowej. Czasami wszystkie rodzaje komórek glejowych określane są wspólnie jako neuroglej.
FUNKCJA: Podobnie do wielkiej różnorodności komórek nerwowych, istnieje duże zróżnicowanie komórek glejowych. Jednak o ile wszystkie neurony pełnią właściwie tę samą funkcję, komórki glejowe mogą pełnić bardzo różne funkcje fizjologiczne. Komórki glejowe współtworzą barierę krew-mózg, pełnią istotną rolę w odżywianiu komórek nerwowych składnikami odżywczymi transportowanymi układem krążenia, syntetyzują wiele istotnych enzymów używanych m.in. do wytwarzania neuroprzekaźników, tworzą osłonki mielinowe aksonów, a nawet pełnią funkcje ochronne. Współczesne badania pokazują również i takie funkcje, jakich dotychczas nikt się nie spodziewał po komórkach glejowych. W dorosłym mózgu ssaków, w tym ludzi, istnieje pewna mała liczba komórek prekursorowych oligodendrocytów, których znaczenie i funkcje pozostają nieznane, aczkolwiek zaobserwowano, że przy działaniu czynników uszkadzających istniejące dotychczas oligodendrocyty, komórki prekursorowe "aktywują się" i rozwijają się z nich nowe oligodendrocyty, chociaż w przypadku poważnych uszkodzeń taki mechanizm nie jest efektywny w "naprawianiu" oligodendrocytów. W 2008 roku odkryto w mózgu szczura, że niektóre z tych komórek prekursorowych zawierają w swojej błonie komórkowej napięciowo zależne kanały sodowe i potasowe, zaś ich błona jest wrażliwa na depolaryzację i otrzymuje synapsy pobudzające i hamujące z aksonów komórek nerwowych. Ta grupa komórek jest w stanie generować potencjał czynnościowy podobnie, jak to czynią komórki nerwowe, podczas gdy komórki glejowe nie posiadają takich zdolności. Wykazano również, że glejowe komórki węchowe mogą stymulować regenerację uszkodzonych aksonów w rdzeniu kręgowym.

a) Mikroglej (ang. microglia, third element) - są to komórki nieneuronalne centralnego układu nerwowego, tkankowo specyficzne, rezydentne makrofagi kontrolujące homeostazę i biorące udział w odpowiedzi immunologicznej.
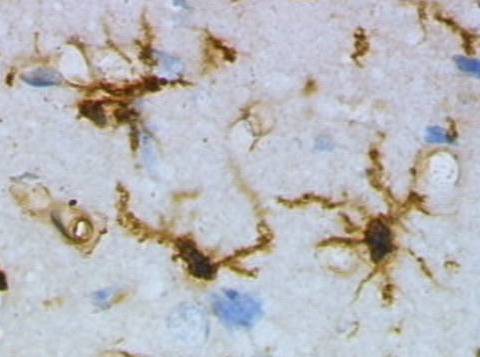 |
| Źródło: Wikipedia. Komórki mikrogleju - forma spoczynkowa lub wczesna faza aktywacji - kora mózgowa szczura barwiona lektynami (kolor brązowy); jądra komórkowe podbarwione hematoksyliną (kolor niebieski). |
HISTORIA: Pierwszymi poważnymi badaniami nad opisem gleju zajął się Santiago Ramon y Cajal (1852-1934) który scharakteryzował trzy podstawowe rodzaje komórek glejowych. Doniósł on o obecności komórek o kształcie gwieździstym - nieświadom tego że są to astrocyty, komórek włóknistych z licznymi wypustkami i komórek protoplazmatycznych występujących licznie w istocie szarej. Jak się okazało później jest to mikroglej. Niezależnie odkryty przez Nissla i Robertsona, dokładniejszy opis zyskał jednak dzięki uczniowi Cajala, Pio del Rio-Hortedze (1882-1945). W wyniku barwienia solami srebra Rio-Hortega wyróżnił specyficzny rodzaj komórek zawierających jądro o wyraźnie wydłużonym kształcie. Scharakteryzowane przez niego komórki mikrogleju zyskały od tej pory miana komórek Hortegi. Następny "krok milowy" w badaniach nad mikroglejem należał znów do Hortegi, który opisywał rolę odkrytego mikrogleju w patologii centralnego układu nerwowego. W późniejszych latach, w wyniku zidentyfikowania komórkowo specyficznych markerów narastały kontrowersje wokół rozwoju embrionalnego jak i pochodzenia mikrogleju. Te i inne wątpliwości rozwiały zaawansowane badania w latach 80. z użyciem markerów lektynowych oraz rozwiniętych technik immunocytochemicznych.
POCHODZENIE: Na temat pochodzenia mikrogleju prowadzono już wiele debat, na podstawie których nawet obecnie trudno jednoznacznie określić rodowód tych komórek. Zdania są podzielone i część badaczy stoi na stanowisku, że źródłem mikrogleju są komórki linii monocytarnej, a więc prekursorów dopatrują się w mezodermie. Dowodem popierającym mezodermalną tezę są badania prowadzone przez Ling (Ling i in. 1980) polegające na znakowaniu monocytów z krwi obwodowej noworodków koloidalnym węglem i późniejszym lokalizowaniu ich w tkance nerwowej mózgu. Niepodważalnym dowodem jest również fakt, że komórki mikrogleju posiadają na swojej powierzchni markery F4/80, Mac-1, ED1, lektyny (GSA I-B4) typowe dla monocytów i makrofagów oraz receptory na Fc, CR3 (Perry i in. 1985). Faktem potwierdzającym założenie mezodermalnej teorii jest również obecność w cytoplazmie mikrogleju elementów lysozymu, oraz takich enzymów jak niespecyficzna esteraza czy peroksydaza.
Zgodnie z mezodermalną teorią pochodzenia komórek mikrogleju monocyty infiltrują parenchymę mózgu we wczesnych okresach życia embrionalnego jako mikroglej amebowaty. Zaznaczyć trzeba że napływ mikrogleju do mózgu jest dodatnio skorelowany z rozwojem unaczynienia tkanki nerwowej co również przemawia za mezodermalnym źródłem mikrogleju (Perry i in. 1985, Miyake i in. 1984, Hurley i in. 1996). Opuszczenie naczyń krwionośnych przez te komórki jest możliwe gdyż bariera krew-mózg tworzona w tym czasie przez astrocyty nie jest jeszcze dokładnie zamknięta w tym stadium życia organizmu. Po przeniknięciu dochodzi do transformacji mikrogleju amebowatego w mikroglej spoczynkowy charakteryzujący się obkurczoną częścią cytoplazmatyczną oraz licznymi rozgałęzieniami (Boya i in. 1991, Fedorolf i in. 1995).
Jako alternatywne źródło mikrogleju podaje się neuroektodermę, z której wywodzą się glioblasty będące prekursorami astrocytów i oligodendrocytów (Fujita i in. 1975, 1980). Jednyzm z dowodów ppopierających teorię ektodermalnego pochodzenia mikrogleju jest fakt, iż mikroglej, jak i komórki progenitorowe 0-2A, z których mogą się rozwinąć oligodendrocyty oraz astrocyty typu drugiego, posiadają jednakowe właściwości histochemiczne. Poza tym zidentyfikowano homologiczne epitopy powierzchniowe zlokalizowane na astrocytach, jak i na mikrogleju, a rozpoznawane przez przeciwciała monoklonalne LN-1 (Dickson i in. 1989). Zgodnie z neuroektodermalną teorią o pochodzeniu mikrogleju zakłada się, że mikroglej wykształcił się jako samodzielna linia z puli komórek macierzystych neurogleju. Faktem przemawiającym za tą teorią jest to, że glioblasty ze strefy okołokomorowej wykształconej z cewki nerwowej są prekursorami komórek Hortegi. Znaczyłoby to, że oligodendrocyty astrocyty i mikroglej są grupami siostrzanymi pochodzącymi z tej samej linii multipotencjalnych komórek neuroektodermalnych (Fujuta i in. 1975, 1980).
Ze względu na brak jednoznacznych i niepodważalnych faktów przemawiających za jedną z opcji pochodzenia mikrogleju proponowane jest alternatywne heterogeniczne źródło (De Groot i in.). Według tego złożenia część mikrogleju zasiedlającego tkankę nerwową jest pochodzenia mezodermalnego, a część neuroektodermalnego. Bez względu na pochodzenie mikrogleju, przyjmuje on w mózgu formy i funkcje charakterystyczne tylko dla tego rodzaju komórek gleju w dużym stopniu zależne od stanu fizjologicznego tkanki.
Funkcja mikrogleju: Mikroglej stanowiący około 5-20% populacji komórek nieneuronalnych w mózgu występuje w warunkach normalnych w formie spoczynkowej. Główne funkcje fizjologiczne mikrogleju sprowadzają się do monitorowania mikrośrodowiska tkanki, usuwania umierających neuronów i reakcji w wyniku stwierdzenia obecności obcego antygenu (Nakajima i in. 1993). W formie nieaktywnej mikroglej posiada liczne i długie rozgałęzienia części cytoplazmatycznej komórki. Ta specyficzna struktura prawdopodobnie ułatwia potencjalną reakcję tych komórek jako specyficznego dla tkanki nerwowej elementu układu immunologicznego.
Charakterystyczna transformacja mikrogleju z formy spoczynkowej do aktywowanej została poznana i opisana przez Rio Hortege prawie wiek temu. W wyniku aktywacji dochodzi do przemian morfologicznych którym towarzyszy: zwiększenie rozmiaru ciała komórki oraz obkurczenie rozgałęzień. Na poziomie molekularnym dochodzi do ekspresji białek adhezyjnych, reorganizacji cytoszkieletu oraz ekspresji elementów kompleksu zgodności tkankowej MHC typu I jak i II (Perry i in. 1987, 1989, Aloisi i in. 2000).
Zaobserwowano, że aktywowanie mikrogleju może odgrywać rolę w ciężkich postaciach zaburzeń psychiatrycznych.
MIKROGLEJ W USZKODZONEJ TKANCE NERWOWEJ: W wyniku uszkodzenia tkanki nerwowej dochodzi w mózgu do odpowiedzi komórkowej i chemicznej otaczających komórek które to reakcje są efektem fizycznego naruszenia integralności tkanki nerwowej, jak i zmian w lokalnym mikrośrodowisku, np. zmiany homeostazy jonowej. Jednocześnie z uszkodzeniem uruchamianych jest wiele procesów mających na celu doprowadzić z jednej strony do ochrony przed czynnikami uszkadzającymi, a z drugiej do naprawy powstałych zniszczeń tkanki (Berkenbosch i in. 1992).
Odpowiedzi komórkowej na czynnik uszkadzający w większości przypadków towarzyszy: proliferacja i migracja komórek mikrogleju i astrocytów, produkcja cytokin prozapalnych, funkcjonalne zmiany w śródbłonku naczyń krwionośnych oraz rekrutacja z krwiobiegu komórek układu immunologicznego w obręb uszkodzonej tkanki (Dong i in. 2001, Aloisi i in. 2001). W uszkodzonych neuronach dochodzi do ekspresji wczesnych genów odpowiedzialnych za stymulację i aktywowanie astrocytów i mikrogleju (Neumann i in. 2001, Raivich i in. 1999). A poprzez przerwaną ciągłość bariery krew-mózg aktywowane zostają monocyty z krwi obwodowej które na wzór mikrogleju ulegają następnie transformacji morfologicznej (Fujuta i in. 1998, Leong i in. 1992, Maxwell i in. 1990).
Aktywacja mikrogleju jaką wywołuje uszkodzenie tkanki związana jest ze wzrostem poziomu czynników o funkcji immunologicznej. W okresie tym może dojść do indukcji ekspresji szerokiej rzeszy receptorów powierzchniowych które przyspieszają odpowiedź immunologiczną. Pośród nich są receptory biorące udział w rozpoznawaniu cząsteczek związanych z patogenem, receptory komplementu (np. CR1, CR3, CR4), receptory cytokin (np. TNFRI, TNFRII, IL-1RI, IL-12R) oraz chemokin (np. CCR2, CCR3, CXCR4, CX3CR1) jak i receptory ułatwiające interakcje z układem immunologicznym jak np. z limfocytami T czy immunoglobulinami (np. Fc RI, RII, RIII) (Aloisi i in. 2001, Perry i in. 1992, Nakajima i in. 1993).
 |
| Źródło: Wikipedia. Oligodendrocyt. |
 |
| Źródło: Wikipedia. Morfologiczne zmiany różnicującego się oligodendrocytu (barwienie na czynnik transkrypcyjny Olig 2, czerwony, oraz CNP-azę, zielony). |
 |
| Źródło: Wikipedia. Oligodendrocyt w pierwszej fazie mielinizacji. Barwienie na czynnik transkrypcyjny Olig2 (czerwony) oraz CNPase (zielony). |
W wyniku uszkodzenia centralnego układu nerwowego, jak również w wyniku chorób neurodegeneracyjnych, dochodzi także do degeneracji i śmierci oligodendrocytów.
Oligodendrocyty eksprymują na swojej powierzchni serie charakterystycznych białek - Nogo-A, MAG, OMgp i inne, które wiązane są silnie z ich hamującymi właściwościami na wzrost zakończeń nerwowych po uszkodzeniu. Teza ta jednak stoi przed wyzwaniem rzuconym przez grupę prof. Schwaba pokazującą nagły i silny wzrost zakończeń nerwowych wzdłuż zmielinizowanych szlaków nerwowych nie tworzących rozgałęzień do sąsiadujących szlaków. Nazwano to z angielskiego guard rai poprzez negatywną stymulację. Co więcej, ostatnie badania in vivo sugeruję, że MAG, komponent mieliny uznawany za inhibitor wzrostu, w rzeczywistości wzmaga wzrost zakończeń nerwowych.
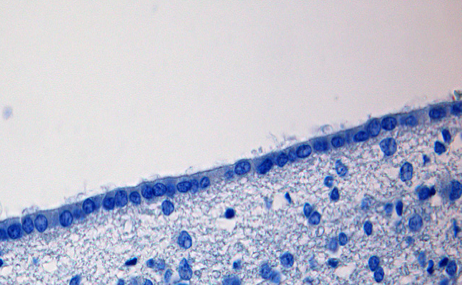 |
| Źródło: Wikipedia. Wyściółka, barwienie H-E. |
Funkcją ependymy wchodzącej w skład bariery krew-mózg jest uczestniczenie w wymianie substancji pomiędzy płynem mózgowo-rdzeniowym a tkanką nerwową.
d) Komórka Schwanna, także lemocyt, lemmocyt - komórka glejowa obwodowego układu nerwowego występująca w zespołach.
 |
| Źródło: Wikipedia. Komórka Schwanna z hodowli komórkowej. |
Osłonka wewnętrzna (mielinowa): we włóknach nerwowych rdzennych (tzw. białych) komórki Schwanna tworzą wzdłuż aksonów osłonkę mielinową ze swojej błony komórkowej przez wielokrotne owinięcie się jednej komórki wokół fragmentu aksonu długości 50 mikrometrów - 1 mm. Osłonka ta spełnia funkcję ochronną dla aksonu, ale przede wszystkim zwiększa tempo przewodzenia impulsów nerwowych (dzięki przewężeniom Ranviera).- Osłonka zewnętrzna (Schwanna): we włóknach bezrdzennych (szarych) osłonka Schwanna (neurolemma, neurylemma) powstaje jedynie przez okrycie aksonu protoplazmą komórki Schwanna ("zatopienie" włókna w cytoplazmie). Jedna osłonka pokrywa kilka z nich. Cytoplazma komórki Schwanna wiąże aksony razem, ale nie pozwala im się dotykać.
 |
| Źródło: Wikimedia. Komórki satelitowe. |
Brak komentarzy:
Prześlij komentarz