

 1) Skandowce – pierwiastki 3 (daw. IIIB lub III pobocznej) grupy układu okresowego. Grupa skandowców, podobnie jak inne grupy należące do bloku d układu okresowego, powinna zawierać cztery pierwiastki, jednak nie ma zgodności co do przypisania pierwiastków z 6 i 7 okresu. Dwa pierwsze pierwiastki, skand (Sc, okres 4) i itr (Y, okres 5), nie budzą kontrowersji, natomiast pozostałe dwa klasyfikowane są różnie. IUPAC zalicza do tej grupy lutet (Lu) i lorens (Lr), według niektórych autorów są to natomiast lantan (La) i aktyn (Ac). Czasem przyjmuje się też, że grupa ta zawiera wszystkie lantanowce i aktynowce (łącznie 32 pierwiastki) lub przeciwnie, zalicza się do niej tylko skand i itr, a wszystkie lantanowce i aktynowce klasyfikuje się osobno. Skand, itr i lantanowce są nazywane łącznie metalami ziem rzadkich.
1) Skandowce – pierwiastki 3 (daw. IIIB lub III pobocznej) grupy układu okresowego. Grupa skandowców, podobnie jak inne grupy należące do bloku d układu okresowego, powinna zawierać cztery pierwiastki, jednak nie ma zgodności co do przypisania pierwiastków z 6 i 7 okresu. Dwa pierwsze pierwiastki, skand (Sc, okres 4) i itr (Y, okres 5), nie budzą kontrowersji, natomiast pozostałe dwa klasyfikowane są różnie. IUPAC zalicza do tej grupy lutet (Lu) i lorens (Lr), według niektórych autorów są to natomiast lantan (La) i aktyn (Ac). Czasem przyjmuje się też, że grupa ta zawiera wszystkie lantanowce i aktynowce (łącznie 32 pierwiastki) lub przeciwnie, zalicza się do niej tylko skand i itr, a wszystkie lantanowce i aktynowce klasyfikuje się osobno. Skand, itr i lantanowce są nazywane łącznie metalami ziem rzadkich.


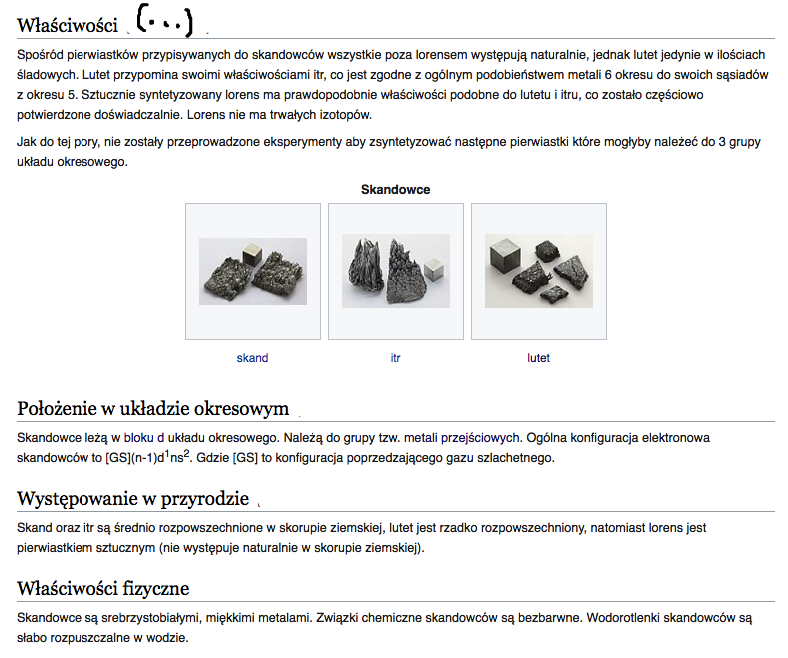
Właściwości chemiczne
Skandowce są bardzo podobne pod względem chemicznym. Charakteryzują się niską elektroujemnością. W związkach są na III stopniu utlenienia. W podwyższonej temperaturze reagują z tlenem, tworząc tlenki. Z wodą tworzą wodorotlenki, których charakter zasadowy rośnie "w dół" grupy. Wodorotlenek skandu jest amfoteryczny.
Otrzymywanie
Skandowce są otrzymywane głównie przez elektrolizę ich stopionych soli.








2) Tytanowce - są to pierwiastki 4 (daw. IVB lub IV pobocznej) grupy układu okresowego. Są to: tytan (Ti), cyrkon (Zr), hafn (Hf) i rutherford (Rf).

Położenie w układzie okresowym: Tytanowce leżą w bloku d układu okresowego. Są typowymi pierwiastkami przejściowymi. Ogólna konfiguracja elektronowa tej grupy to [GS](n-1)d2ns2, gdzie [GS] to konfiguracja poprzedzającego gazu szlachetnego.
Występowanie w przyrodzie: Występują w skorupie ziemskiej w stosunkowo dużych ilościach: tytan stanowi około 0,42%, cyrkon - 0,02%, a hafn - 0,005%. Minerały tytanu i cyrkonu składają się głównie z tlenków tych metali. Rutherford został otrzymany sztucznie i nie występuje na Ziemi.
Właściwości fizyczne: Tytan, cyrkon i hafn są metalami srebrzystobiałymi, wytrzymałymi mechanicznie i trudno topliwymi. Właściwości rutherfordu obecnie nie są jeszcze bliżej znane. Tytanowce na powietrzu ulegają pasywacji. Tlenki tytanowców na IV stopniu utlenienia są trudno topliwe, a tlenek cyrkonu ZrO2jest jednym z najbardziej ogniotrwałych materiałów jakie dotychczas poznano.
Właściwości chemiczne: Są reaktywne w wysokich temperaturach. W związkach występują najczęściej na IV stopniu utlenienia, rzadziej na II czy III. Tlenki są amfoteryczne z przewagą cech kwasowych. Halogenki nie mają charakteru soli, łatwo ulegają hydrolizie, sublimują i są lotne.
Otrzymywanie: Tytanowce otrzymuje się przez redukcję ich halogenków metalami alkalicznymi.






















































































Związki tytanu:























Związki cyrkonu:


















































































Związki niobu:
















Związki tantalu:





4) Chromowce:

























Związki molibdenu:





































Związki technetu:













6) Żelazowce - są to pierwiastki chemiczne znajdujące się w grupie 8 układu okresowego (dawniej zaliczane do VIII grupy pobocznej, której pierwszą triadę również nazywano żelazowcami) - są to żelazo (Fe), ruten (Ru), osm (Os) i has (Hs).





























7) Kobaltowce - pierwiastki chemiczne znajdujące się w 9 grupie układu okresowego (dawniej zaliczane do VIII grupy pobocznej) - są to kobalt (Co), rod (Rh), iryd (Ir) i meitner (Mt).

 |
| Źródło: pse-mendelejew.de - Kobalt. |
































Źródło: praca pt. "Kobalt w środowisku przyrodniczym i antropogenicznym" - autor: Andrzej Greinert.

























































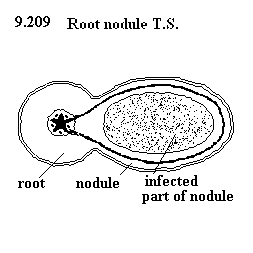 |
| Źródło: uq.edu.au - Rood nodule - Brodawka korzeniowa - schemat. |
 |
| Źródło: Evolution - 2007 - Cold Spring Harbor Laboratory Press. Figure 6.26 - Symbiosis of Rhizobium bacteria with legumes. |







Nawiązanie symbiozy
Nawiązanie symbiozy pomiędzy rizobiami a roślinami motylkowymi ma miejsce w warunkach ograniczonej dostępności azotu. Proces ten przebiega na zasadzie wzajemnej wymiany określonej sekwencji związków sygnalnych inicjujących brodawkowanie poprzez aktywację specyficznych genów roślinnych i bakteryjnych.
Bakterie w odpowiedzi na roślinne struktury i mechanizmy obronne, wykształciły szereg przeciwdziałających im strategii. Jednocześnie korzyści jakie przynosi roślinom symbioza z diazotrofami przyczynia się do wytworzenia przez nie mechanizmów ułatwiających infekcję zgodnym, tj. zdolnym do nawiązania efektywnej symbiozy mikrosymbiontom.
Istotną rolę w procesie infekcji odgrywają wydzielane przez roślinę flawonoidy. Wywołują one dodatnią chemotaksję zgodnych, czyli zdolnych do nawiązania efektywnej symbiozy rizobiów. Związki te są także induktorami bakteryjnych genów nod (ang. nodulation) odpowiadających za brodawkowanie (nodulację) roślin. Zdolność do ich aktywacji zależy od gatunku rośliny. U Rhizobium ulegają aktywacji pod wpływem wydzielanych przez rośliny strefy umiarkowanej (np. koniczyna, lucerna, wyka, groch) flawonów i flawononów np. luteoliny, zaś u Bradyrhizobiumpod wpływem izoflawonów np. daidzeiny, wydzielanych przez tropikalne rośliny motylkowate (np. soja). Geny nod rizobiów zorganizowane są w operony zlokalizowane na chromosomie lub na jednym z tzw. plazmidów symbiotycznych Sym. Wspólne dla wszystkich rizobiów geny com nod (ang. common nodulation genes) – nodABC, o dużej konserwatywności sekwencji służą do syntezy szkieletu głównego czynników Nod, zaś geny hsn nod (ang. host specific nodulation genes) zależnie od gatunku bakterii występują w różnych kombinacjach i określają specyficzność wobec gospodarza. Ekspresję genów nod indukuje białko regulatorowe NodD (m. cz. ok. 35kDa). Gen nodD jest jedynym genem nod ulegającym konstytutywnej ekspresji, zarówno u bakterii wolno żyjących, jak i będących w symbiozie z roślinami. Przypuszcza się, że dany gatunek rizobiów syntetyzuje określone białko NodD, aktywujące transkrypcję genów nod tylko po związaniu z określonym flawonoidem, co determinuje specyficzność względem gospodarza. Ta specyficzność bakterii wobec różnych aktywatorów może wynikać z różnic w wiązaniu, transporcie i oddziaływaniach z białkami regulatorowymi.

Rys.1. Wymiana cząstek sygnałowych pomiędzy symbiontami i odpowiedzi rośliny na przykładzie oddziaływań Rhizobium meliloti i lucerny. 1-deformacja włośników; 2-tworzenie nici infekcyjnej; 3-podziały komórkowe prowadzące do wytworzenia brodawki
Tzw. czynniki Nod (chitolipooligosacharydy; CLOS ), będące produktami genów nod, to morfogeny, które powodują zwijanie się włośników korzeniowych, tworzenie nici infekcyjnej i powstawanie w korze korzenia nowych merystemów, dając początek brodawkom korzeniowym.
Roślinne bakteriostatyczne izoflawonoidowe fitoaleksyny, udaremniają kontakt mikrosymbiontów z ich komórkami, jednak bakterie zgodne wobec danej rośliny w przeciwieństwie do innych mikroorganizmów patogennych są na nie mniej wrażliwe. Kontakt fitoaleksyn z rizobiami ograniczają przylegające do powierzchni komórek bakteryjnych cykliczne beta-glukany, a wydzielane do otoczenia hamują ich produkcję . W ochronie rizobiów przed działaniem izoflawonoidów biorą udział składniki ich struktur powierzchniowych - lipopolisacharydy (LPS) i egzopolisacharydy (EPS).
Rośliny wydzielają również enzymy, takie jak chitynazy i glukanazy, hydrolizujące ściany mikroorganizmów patogennych, oraz związki niskocząsteczkowe, np. cukry i kwasy organiczne służące bakteriom jako substancje odżywcze. Chitynazy wydzielane przez rośliny hydrolizują syntetyzowane przez bakterie czynniki Nod, czemu zapobiegają chemiczne modyfikacje tych czynników wykluczając je z grupy substratów chitynaz. W wydzielinach korzeni obecne są też oddziałujące z EPS - pektyny, powodujące agregację bakterii w sąsiedztwie włośników korzeniowych. Również mikroorganizmy mogą syntetyzować lektyny, które specyficznie wiążą cukry powierzchni komórek odpowiedniej rośliny. Po połączeniu symbiontów z udziałem lektyn i polisacharydów następuje trwała adhezja bakterii do ścian komórek epidermy korzeni za pomocą syntetyzowanych przez rizobia mikrowłókien celulozowych (R. leguminosarum) lub/i białkowych fimbrii (Bradyrhizobium japonicum).
Czynniki Nod aktywują w roślinie tzw. geny nodulinowe, które w zależności od czasu ich ekspresji dzieli się na wczesne - ENOD (ang. early nodulin genes) - wyrażane przed rozpoczęciem wiązania azotu i późne - LNOD (ang. late nodulin genes) - podczas, lub po wiązaniu azotu. Wczesne noduliny biorą udział w infekcji bakteryjnej (deformacja włośników, tworzenie nici infekcyjnej) oraz morfogenezie brodawek (podziały komórkowe, tworzenie zawiązków brodawek). Późne noduliny to białka niezbędne do funkcjonowania brodawki (np. leghemoglobina), enzymy lub podjednostki enzymów uczestniczących w metabolizmie oraz transporcie azotu i węgla, niektóre białka błony peribakteroidalnej. Część białek LNOD znajduje się poza brodawką i ulega ekspresji nie tylko podczas symbiozy.
Po adhezji rozpoczyna się degradacja roślinnych ścian komórkowych przez związane ze ścianami bakterii pektynazy i celulazy, co umożliwia wniknięcie rizobiów do wnętrza włośników. Bakterie przemieszczają się w kierunku zawiązków brodawek poprzez nici infekcyjne. W ich tworzeniu biorą udział roślinne białka strukturalne i lektyny oraz bakteryjny EPS i łańcuch o-swoisty LPS. W obrębie matrix nici infekcyjnej bakterie namnażają się, a po dotarciu do brodawki uwalniane są do cytoplazmy komórek roślinnych. Rizobia na drodze endocytozy zostają otoczone błonami peribakteroidalnymi pochodzenia roślinnego, stanowiącymi powierzchnię wymiany metabolitów i sygnałów pomiędzy mikrosymbiontem a komórką roślinną. Tak powstają symbiosomy, w obrębie których bakterie przekształcają się w zdolne o wiązania azotu atmosferycznego bakteroidy, czyli zdeformowane komórki bakterii o objętości dzisięcio, a nawet trzydziestokrotnie większej niż rizobia.
Informacja genetyczna bakteroidów w postaci genów nif warunkuje syntezę enzymu nitrogenazy. Tzw. kompleks nitrogenazy złożony z nitrogenazy i reduktazy nitrogenazowej znajduje się w ich cytoplazmie i jest wrażliwy na tlen. Reduktaza nitrogenazowa to kodowany przez gen nich homodimer metaloproteiny Fe o m. cz. ok. 60 kDa. Nitrogenazę, która jest tetramerem metaloproteiny Mo-Fe (o m. cz. ok. 240 kDa kodują geny Nid i nifK. Reduktaza nitrogenazowa dostarcza siły redukcyjnej potrzebnej do wiązania azotu, co wymaga zużycia 16 moli ATP na 1 mol N2. Nitrogenaza przenosi uwolnione przez reduktazę nitrogenazową elektrony na N2, czemu towarzyszy redukcja H+ do H2. U niektórych diazotrofów, wodór powstający obok NH3, tworzy wodę dzięki obecności hydrogenazy, redukując tym samym tlen zagrażający aktywności nitrogenazy. Pozwala to również odzyskać część energii zużytej na wytworzenie wodoru, co przedstawia reakcja:
N2 + 8H+ + 16ATP + 8e- → 2NH3 + H2 + 16ADP + 16Pi

Warunki mikroaerofilne, niezbędne do aktywności nitrogenazy, zapewnia przede wszystkim wiążące tlen białko - leghemoglobina (Lb), będąca hemoproteiną o wysokim powinowactwie do tlenu oraz znacznej homologii i podobieństwie strukturalnym do zwierzęcej hemoglobiny. Grupa prostetyczna leghemoglobiny - protohem, jest syntetyzowana przez bakteroidy, zaś synteza części białkowej odbywa się w komórkach roślinnych. Leghemoglobina obecna jest w cytoplazmie komórek roślinnych, nie występuje natomiast w przestrzeni perybakteroidalnej. Nadaje tkance brodawki zawierającej efektywnie wiążące azot bakteroidy barwę czerwoną. Właściwości leghemoglobiny zapewniają bakteroidom wystarczającą ilość tlenu potrzebną do procesów energetycznych, nie przekraczając takiej, która hamuje wiązanie azotu. Dyfuzję gazów przez ściany brodawki ogranicza dodatkowo wysycenie białkami bogatymi w prolinę ścian nie zakażonych komórek, które otaczają tkanki zasiedlone przez rizobia.





Regulacja brodawkowania Wielorakie mechanizmy regulacyjne wpływają na proces brodawkowania (nodulacji). Istnieje hipoteza, że roślinne hormony gł. kwas abscysynowy (ABA), auksyny, cytokininy, etylen i kwas giberelinowy (GA3) regulują rozwój brodawek. Ich proporcja zmieniana jest przez czynniki Nod. Na kształtowanie się brodawek wpływają też hormony peptydowe, tlenek azotu (NO), aktywne/reaktywne formy tlenu (AOS/ROS), kwas jasmonowy (JA), kwas salicylowy (SA), urydyna, flawonoidy i same czynniki Nod. Różne cząstki sygnalne wymieniane pomiędzy rośliną a infekującymi ją bakteriami regulują inicjację, różnicowanie, funkcjonowanie oraz liczbę brodawek.
Co najmniej dwa szlaki regulują liczbę brodawek:
- miejscowa regulacja infekcji we wrażliwej na nią strefie korzenia;
- autoregulacja poprzez ujemne sprzężenie zwrotne, gdzie merystemy istniejących już brodawek wyzwalają sygnały hamujące powstawanie kolejnych.
Ze względu na znaczną energochłonność procesu wiązania azotu, liczba brodawek musi być ściśle dostosowana do wydajności fotosyntetycznej rośliny. Autoregulacja sprawowana jest przez czynnik pochodzący prawdopodobnie z liści, poruszający się w kierunku korzeni. Może on być łącznikiem pomiędzy inicjacją brodawkowania a aktywnością fotosyntetyczną gospodarza.
Kwas abscysynowy (ABA) jest inhibitorem rozwoju brodawek, hamującym stymulowane cytokininami podziały komórek kory korzenia. Na nodulację wpływa proporcja ABA:cytokoniny, istnieje więc interakcja pomiędzy „sygnalingiem” tych hormonów. Pewien model zakłada, że zakażenie indukuje początkowy spadek ilości ABA w ksylemie, po czym oba hormony translokowane są do liści gdzie następuje synteza ABA powracającego poprzez floem do korzeni i hamującego tworzenie kolejnych brodawek. Wzrost ilości ABA w brodawkach może indukować gromadzenie się w nich cukrów, zatem drugą jego funkcją w procesie nodulacji jest przydzielanie (alokacja) brodawkom (organom "sink") fotoasymilatów - źródła energii potrzebnej do wzrostu, rozwoju, oddychania i wiązania azotu przez rizobia. ABA powoduje wzrost akumulacji mRNA lipooksygenazy (LOX), której aktywność w wierzchołkach rozwijających się, ale nie w dojrzałych brodawkach sugeruje, że uczestniczy w ich wzroście i rozwoju. W regionie tym podwyższona jest też ilość ABA. Poza dwojakim wpływem ABA na nodulację tj. negatywnej regulacji ich ilości a pozytywnej wzrostu i rozwoju pojedynczych brodawek, istnieje trzecia jego funkcja. Początkowo wysoki poziom ABA, związany z regulacją ich wzrostu i liczby, osiąga na etapie wiązania azotu plato, po czym znów wzrasta. Wiąże się to z udziałem ABA w starzeniu się brodawek.
Auksyny syntetyzowane głównie w pędach są aktywnie transportowane do korzeni. Podczas nodulacji uczestniczą w podziałach i różnicowaniu się komórek oraz w tworzeniu merystemów. Indukują ekspresję genów cyklu komórkowego potrzebnych do aktywacji podziałów komórkowych. Dlatego podwyższony ich poziom poprzedza tworzenie brodawek i utrzymuje się we wczesnych etapach ich rozwoju, a obniża się w dojrzałych brodawkach. W wielu gatunkach roślin poziom auksyn w brodawkach przewyższa ich ilość w korzeniach. Stosunek auksyn do cytokinin w korzeniach odpowiada za inicjację podziałów komórek kory i powstawanie brodawek oraz regulację ich liczby. Przypuszcza się, że rizobia mogą manipulować poziomem auksyn u roślin hamując ich transport, co zmniejsza ich ilość względem cytokinin i promuje brodawkowanie. Poziom auksyn może być też regulowany poprzez kontrolę ilości ich transporterów. Syntetyczne inhibitory transportu auksyn indukują wytwarzanie pseudobrodawek oraz ekspresję genów nodulin (ENOD 12,40,2). Składniki roślinne tj. etylen, cytokininy, flawonoidy, mogą hamować transport auksyn oraz regulować aktywność rozkładających je enzymów np. peroksydazy, oksydazy kwasu indolilo-3-octowego. Podwyższona aktywność peroksydazy w dojrzałych brodawkach P. vulgaris ma prawdopodobnie ograniczyć w nich wzrost poziomu auksyn. Niektóre flawonoidy hamują rozkład auksyn przez w/w enzymy a inne go stymulują. Wskazuje to na lokalną regulację ich poziomu.
Cytokininy stymulują inicjowane przez rizobia podziały komórek kory korzenia. Fitohormony te, podobnie jak bakteryjne czynniki Nod, indukują też depozycję amyloplastów i ekspresję niektórych genów, np. enod40. Wskazuje to na istnienie w korze wewnętrznej elementów wspólnych dla ich szlaków sygnałowych. Czynniki Nod wywołują prawdopodobnie zmiany w syntezie cytokinin, ich obrocie lub wrażliwości na nie komórek korzenia. Poziom cytokinin w korzeniu zmieniany jest też przez rizobia i jest niższy w porównaniu z brodawkami. Jest też wyższy w młodych (aktywnych mitotycznie) i rozwijających się brodawkach, szczególnie w ich merystemach a spada wraz z osiągnięciem przez nie dojrzałości. Rola cytokinin w nodulacji obejmuje aktywację cyklu komórkowego i związanych z nim genów. Mogą też brać udział w aktywacji części genów ENOD. Zbadano, że część genów nodulin lucerny regulowanych przez cytokininy, indukowanych jest też przez auksyny, co sugeruje częściowe nakładanie się tych szlaków regulacyjnych. Przypuszczalną rolą cytokinin jest też skierowywanie węglowodanów do rozwijających się brodawek. Sądzi się, że nie same cytokininy lecz stosunek cytokininy:auksyny pełni istotną rolę w nodulacji.
Poziom etylenu wzrasta lokalnie w korzeniach lucerny, wyki i soi zakażonych rizobiami. Wiąże się to z początkiem odpowiedzi obronnej rośliny. Zastosowanie inhibitorów syntezy lub percepcji etylenu (np. Ag2+) powoduje wzrost ilości brodawek. Zakłada się, że etylen indukuje roślinne chitynazy niszczące czynniki Nod. W przypadku grochu (pea) etylen blokuje raczej wejście rizobiów do kory korzenia a nie samą infekcję. Negatywna regulacja etapu infekcji przez etylen ma miejsce u M. truncatula. Etylen nie jest więc zaangażowany w systemową/układową autoregulację nodulacji. Bierze raczej udział w regulacji częstości infekcji działając miejscowo w podatnych na nią strefach korzenia. Rola etylenu w nodulacji może zależeć od infekujących roślinę rizobiów. W grochu poprzez oddziaływanie na cytoszkielet, etylen może być zaangażowany w tworzenie nici preinfekcyjnej i infekcyjnej oraz prawdopodobnie w określanie położenia zawiązków brodawek. Wykazano, że etylen zmienia fenotyp brodawek Sesbania rostrata, która w przypadku jego braku tworzy brodawki niezdeterminowane, a w obecności - zdeterminowane. Sugeruje to udział etylenu w różnicowaniu merystemu. Rhizobitoksyna produkowana przez Bradyrhizobium elkami działa jako inhibitor syntezy etylenu i wzmacnia nodulację siratro (Macroptilium atropurpureum).
Gibereliny w brodawkach wielu roślin strączkowych występują w większych ilościach niż w sąsiednich tkankach korzenia. Dodanie kwasu giberelinowego (GA3) do ekstraktu brodawek L. luteus stymuluje produkcję kwasu indolilo-3-octowego (IAA - auksyna), sugeruje się istnienie pomiędzy nimi sygnalingu. GA3 może zwiększać bioprodukcję lub zmniejszać metabolizm IAA. Zatem nie same gibereliny, co stosunek gibereliny:IAA istotny jest dla nodulacji. Różne szczepy rizobiów zdolne są do syntezy giberelin, mogą więc regulować ich poziom w brodawkach. Istnieje hipoteza, że poza szczepem bakteryjnym także azotan, ABA, a nawet gatunek rośliny wpływa na stężenie giberelin w brodawkach. Pozytywną regulację podziałów komórek pericyklu, koniecznych do wytworzenia brodawki, sprawowaną przez gibereliny hamuje azotan. Wzrost stężenia giberelin może nie być konieczny do tworzenia brodawek np. przez P. vulgaris. Możliwe więc, że wzrost ilości giberelin w brodawkach jest jedynie konsekwencją wysokich poziomów IAA. Gibereliny mogą też działać jako sygnały do hydrolizy skrobi w brodawkach w celu dostarczenia rizobiom substratów oddechowych. Hormony te promują produkcję alfa-amylazy. Istnieje związek pomiędzy giberelinami a alfa-amylazą w reakcjach hydrolizy u różnych gatunków grzybów. Możliwe więc, że bakterie odpowiedzialne są za regulację poziomów giberelin w brodawkach, mających znaczenie w dostarczaniu im substancji odżywczych. Podwyższone poziomy giberelin w brodawkach skorelowane są ze wzrostem liczby i długości międzywęźli oraz ze wzrostem długości ogonków liściowych P. lunatus i Dolichos chinensis(wspięga/fasolnik). Gibereliny mogą zatem przynosić korzyści obu symbiontom, zwiększając rozmiary rośliny co podnosi jej możliwości fotosyntetyczne. Daje to więcej fotoasymilatów zarówno roślinie, jak i brodawkom do ich wzrostu i funkcjonowania.
Źródło: e-biotechnologia.pl - "Symbioza bakterii z roślinami", autor: Adam Kuzdraliński.

"The effects of cobalt deficiency on nodule formation and function in sweet lupin (Lupinus angustifolius L. cv. Unicrop) were studied in cobalt-deficient Lancelin sand in the glasshouse. Bacteroid densities in cobalt-deficient nodules were lower than in normal nodules. Recovery from cobalt deficiency in inoculated treatments was associated with increases in bacteroid density and cobalt accumulation in lateral nodules. Such changes did not occur in treatments infected with rhizobia from the soil. Acetylene-reducing activity of cobalt-deficient plants was not initiated until plants were nearly 6 weeks old, at which time cobalt-treated plants were at their peak of activity. Specific activities of cobalt-deficient nodules remained very low even when nitrogenase did develop. Their large mass of nodules allowed cobalt-deficient plants to reach 20 to 50% of the normal activity per plant, but specific activities were only 5 to 13% of peak activities in cobalt-treated nodules. Nodule bacteroid content and leghaemoglobin content were linearly related to cobalamin content, each with a single relationship. Plotting acetylene-reducing activity against cobalamin content or leghaemoglobin content generated two different linear response curves in each case; the slopes of the lines were different, depending on the presence or absence of cobalt. It is suggested that there may be a function in N2 fixation in legume nodules for a non-cobalamin form of cobalt" --> Źródło: "Cobalt and Nitrogen Fixation in Lupinus angustifolius L. II. Nodule Formation and Function" - M. J. Dilworth, A. D. Robson and D. L. Chatel.
Zagadnienia związane z kobaltem:




Związki kobaltu:








Związki rodu:























Związki irydu:







8) Niklowce - są to pierwiastki chemiczne znajdujące się w 10 grupie układu okresowego (dawniej zaliczane do VIII grupy pobocznej). Należą one do grupy metali przejściowych. Są nimi: nikiel (Ni), pallad (Pd), platyna (Pt) i darmsztadt (Ds).













































Terminy związane z niklem:



















































Związki niklu:


































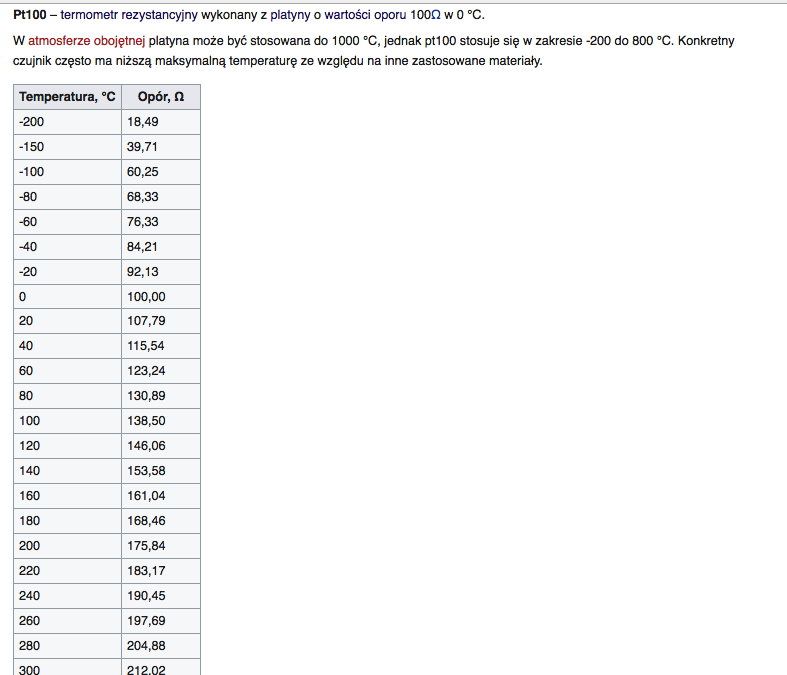














Związki platyny:
























9) Miedziowce - pierwiastki chemiczne znajdujące się w 11 (daw. IB lub I pobocznej) grupie układu okresowego. Należą one do grupy metali przejściowych. Są to: miedź (Cu), srebro (Ag), złoto (Au) i roentgen (Rg).

































































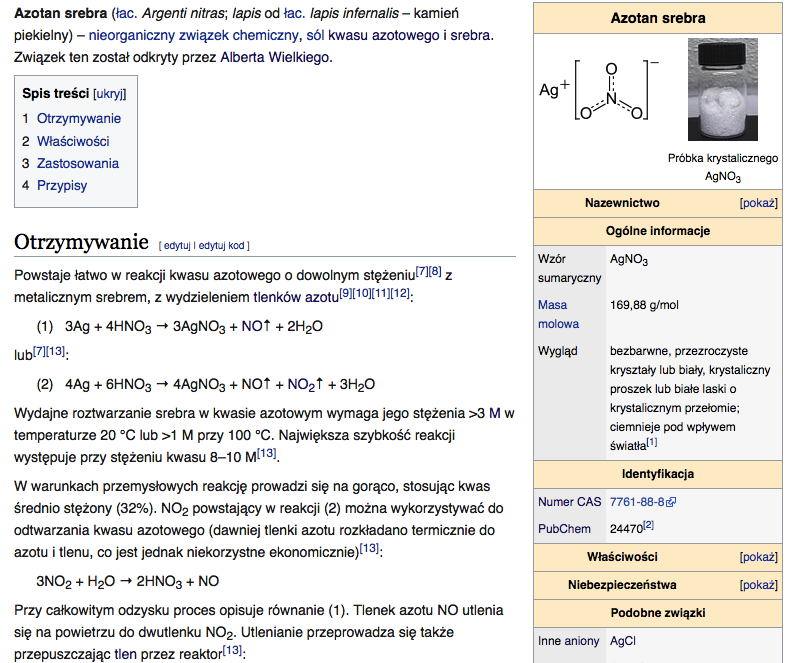








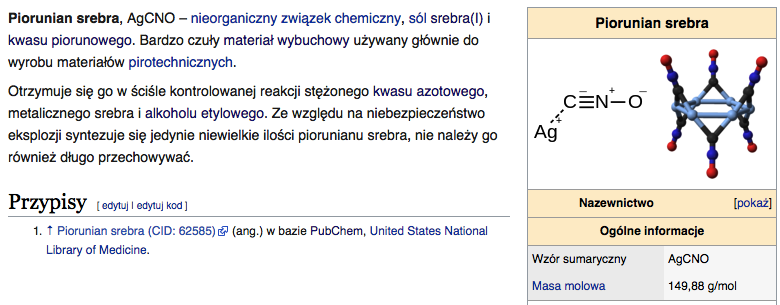


























































































































































10) Cynkowce - są to pierwiastki chemiczne należące do 12 (daw. IIB lub iI pobocznej) grupy układu okresowego. Do cynkowców należą następujące metale: cynk (Zn), kadm (Cd), rtęć (Hg) i kopernik (Cn). Na powłoce walencyjnej cynkowców znajdują się dwa elektrony walencyjne.























































































Cynk w centrach aktywnych enzymów:














Związki kadmu:















































 Związki artęci:
Związki artęci:
- Odczynnik Nesslera,
- Tlenek rtęci (II):

- Zasada Miltona
















































- Odczynnik Nesslera,
- Tlenek rtęci (II):

- Zasada Miltona













- Dimetylortęć,
- Metylortęć,














































































Brak komentarzy:
Prześlij komentarz