
Informacja wstępna: Wirusologia jest działem mikrobiologii zajmującym się badaniem wirusów, ich systematyką, budową, właściwościami antygenowymi i chorobotwórczymi. Wirusologia opracowuje także metody izolacji, oczyszczania, namnażania i zwalczania wirusów. W celach badawczych korzysta się w niej z mikroskopu elektronowego.
Dyskusja z dr hab. n. med. Anną Piekarską na temat szczepień i współczesnych problemów wirusologii (czyli trochę praktycznych informacji na temat tego jak powinniśmy i jak próbujemy radzić sobie z zakażeniami wirusowymi):
Historia badań nad wirusami:

Prowadził badania nad rakotwórczą rolą niektórych kancerogennych wirusów, za co został wyróżniony Nagrodą Nobla w 1966 roku (jeden z najstarszych laureatów w historii), razem z Charlesem Hugginsem. W roku 1911 wykrył pierwsze takie wirusy, zdolne do samodzielnej replikacji, wywołujące u kur mięsaka (nazywane wirusami mięsaka Rousa – Rous sarcoma virus). Udowodnił doświadczalnie, że mięsak można przenosić nie tylko drogą zwykłego przeszczepienia, ale również stosując bezkomórkowy przesącz i jest wywołany przez wirus. Opracowane przez niego techniki przechowywania krwiprzyczyniły się do powstania pierwszych banków krwi.

Punkty - zagadnienia omówione w tym poście:
Dyskusja z dr hab. n. med. Anną Piekarską na temat szczepień i współczesnych problemów wirusologii (czyli trochę praktycznych informacji na temat tego jak powinniśmy i jak próbujemy radzić sobie z zakażeniami wirusowymi):
Historia badań nad wirusami:
- 1892r. - Dmitrij Iwanowski - odkrył, że chorobę mozaikową tytoniu i ziemniaka wywołują zarazki przesiąkające przez filtry bakteryjne.
- 1910r. - Peyton Rous - stwierdził, że wirusy wywołują u zwierząt objawy rakotwórcze.
 |
| Źródło: nobelprize.org - Francis Peyton Rous. |
- 1917r. - Félix Hubert d'Herelle odkrył wirusy niszczące bakterie tzw. bakteriofagi + jest on prekursorem terapii fagowej. Więcej informacji na temat badań prowadzonych przez d'Herelle znajdziesz w moi filmie na temat prionów i wiroidów.
- 1931r. - Ernst Ruska - skonstruował pierwszy mikroskop elektronowy. Link do wykładu noblowskiego "The Development of the Electron Microscope and of Electron Microscopy" - znajdziesz: tutaj.
 |
| Źródło: thefamouspeople.com - Ernst Ruska. |
1935r. - Wendell Meredith Stanley - doprowadził do postaci krystalicznej wirusa mozaiki tytoniowej,- 1949r. - Alfred Day Hershey odkrył u bakteriofagów rekombinację.
- 1952r. - Alfred Day Hershey i Martha Chase udowodnili na przykładzie bakteriofagów, że DNA jest podstawowym nośnikiem informacji genetycznej.
Punkty - zagadnienia omówione w tym poście:
1) Wiroidy i priony,
2) Budowa i formy wirusów. Definicje wirusów.
3) Cykle infekcyjne wirusów,
4) Dualistyczna natura wirusów,
5) Choroby wirusowe i szczepienia ochronne.
1) Wiroidy i priony:
Zachęcam do zapoznania się z filmem mojego autorstwa na temat prionów i wiroidów:
Zachęcam do zapoznania się z filmem mojego autorstwa na temat prionów i wiroidów:
Polecam Wam również filmy z kanału YouTube ADAMED SmartUP:
- Czym są choroby prionowe? / What are prion diseases? - Takao Ishikawa,
- Czy priony zawsze są szkodliwe? / Are prion always harmful? - Takao Ishikawa
2) Budowa i formy wirusów. Definicje wirusów.
 |
| Źródło: Wikipedia (EN) - André Michel Lwoff. |
Wirus (łac. Virus - trucizna, jad) - są to skomplikowane cząsteczki organiczne, nieposiadające struktury komórkowej, zbudowane z białek oraz kwasów nukleinowych. Zawierają one materiał genetyczny w postaci RNA (wirusy RNA) lub DNA (wirusy DNA), wykazują jednak zarówno cechy komórkowych organizmów żywych, jak i materii nieożywionej. Wg definicji Andrego Lwoffa, wirus to "zakaźny", potencjalnie patogenny nukleoproteid, istniejący tylko pod postacią jednego kwasu nukleinowego, który reprodukuje materiał genetyczny. Jest niezdolny do podziałów poza komórką i zazwyczaj nie posiada enzymów (a zatem nie wykazuje metabolizmu).
Czy wirusy są żywe? To, czy należy uznać wirusy za organizmy żywe, zależy od przyjętej definicji życia. Wirusy są zazwyczaj uważane za żywe w funkcjonalnych definicjach życia, nie zaś w definicjach strukturalnych. Funkcjonalne definiowanie życia polega na ustaleniu listy aksjomatów nieodwołujących się do jego struktury, które musi spełniać każdy organizm, żeby można było go uznać za żywy. Według jednej z możliwych aksjomatyzacji (choć nie wszystkie one znaczą mniej więcej to samo) musi on:
- być zdolny do powielania się,
- wykazywać dziedziczną zmienność, wpływającą na zdolność i możliwości rozmnożenia się, czyli być zdolnym do ewolucji.
Priony, choć potrafią się powielać, nie posiadają istotnych cech dziedzicznych, zatem nie są żywe. Wirusy biologiczne, hipotetyczne przedkomórkowe życie na Ziemi, ewentualne podobne formy mogące istnieć na innych planetach - są według tejże definicji żywe.
Strukturalne definiowanie życia polega na polega na ustaleniu listy kryteriów odwołujących się do struktury organizmu. Są to m.in.:
- rozmnażanie,
- wzrost,
- metabolizm,
- budowa komórkowa, z rybosomami i innymi organellami,
- materiał genetyczny przechowywany w postaci kwasów nukleinowych,
- występowanie białek i kwasów nukleinowych.
W przeciwieństwie do definicji funkcjonalnej, nie ma tu ustalonego zbioru warunków; w większości takich zestawów wirusy nie spełniają jednak przynajmniej jednego warunku.
Skąd wirusy pochodzą i czy są żywe? --> Kwestia sporna: Istnieją "skamieniałości" wirusów, które zachowały się w DNA zainfekowanych przez nie gospodarzy, włączając w to Ciebie. Ten ślad "molekularnej skamieliny" (ang. molecular fossil) może pomóc nam zrozumieć, skąd pochodzą wirusy, jak ewoluowały, a może nawet pomóc nam stawić czoła największym pytaniom takim jak np. to, czy wirusy są żywe?
 |
| Źródło: nt.interia.pl - Pandoravirus salinus. |
Podstawowe właściwości wirusów: Wirusy posiadają małe rozmiary. Zdecydowana większość przedostaje się przez filtry mikrobiologiczne zatrzymujące bakterie (są od nich mniejsze). Jednym z największych wirusów są pandorawirusy (Pandorawirusy to rodzaj wirusów DNA. W chwili odkrycia w 2013r. tylko 7% ich genów było podobne do dotychczas znanych genów, z innych organizmów, stąd odkrywcy spodziewający się zaskakujących odkryć, nazwali je od puszki Pandory. Ich DNA zawiera ok. 2 mln zasad. Są to jedne z największych wirusów o rozmiarach 1x0,5 mikrometrów, większe nie tylko od bakterii, ale także od wielu pasożytniczych komórek eukariotycznych; mniejsze jednak od odkrytego nieco później wirusa Pithovirus sibericum.
 |
| Źródło: nature.com - Julia Bartoli & Chantal Abergel - Information Génomique et Structurale - Pithovirus sibericum. |
Pithovirus sibericum został odkryty w 30 000-letniej próbce wiecznej zmarzliny syberyjskiej przez grupę kierowaną przez Chantal Abergel i Jeana-Michela Claverie z Aix-Marseille University. Wyizolowano go z warstwy położonej 30 metrów pod powierzchnią ziemi, pochodzącej z późnego plejstocenu. Stwierdzono, że próbki zebrane w 2000 r. zawierały ameby. Ameby zaczęły ginąć, a po zbadaniu stwierdzono w nich obecność wirusów o niespotykanych dotąd rozmiarach. Chociaż wirus jest nieszkodliwy dla ludzi, jego żywotność po zamrożeniu na przestrzeni tysiącleci wzbudziła obawy, że globalne zmiany klimatu i operacje wiercenia tundry mogą doprowadzić do uwolnienia patogennych wirusów. Jednak inni naukowcy uważają, że ten scenariusz nie stanowi realnego zagrożenia.
Pithovirus sibericum jest gatunkiem gigantycznego wursa o rozmiarach 1,5x0,5 mikrometra, który infekuje ameby. Jest to dwuniciowy wirus DNA. Został odkryty w 2014r. w 30 000 - letnim rdzeniu lodowym [Rdzeń lodowy – próbka rdzeniowa z wieloletniej akumulacji śniegu i lodu, które zrekrystalizowały i uwięziły pęcherzyki powietrza oraz inne osady (np. pył wulkaniczny) pochodzące z różnych okresów. Ze składników rdzenia lodowego, szczególnie obecności izotopówwodoru i tlenu, możliwe jest odczytanie informacji o klimacie w przeszłości, m.in. temperatury, opadów, zawartości gazów i aerozoli atmosferycznych], zebranym z wiecznej zmarzliny na Syberii w Rosji. Nazwa "Pithovirus" jest odniesieniem do dużych pojemników do przechowywania w starożytnej Grecji, znanej jako pithoi. Pithovirus sibericum jest o 50% większy niż największe znane wcześniej pandorawirusy i jest większy niż Ostreococcus, najmniejsza komórka eukariotyczna. Pithovirus ma grubą, owalną ścianę z otworem na jednym końcu. Wewnętrznie jego struktura przypomina plaster miodu. Genom P. sibericum zawiera 467 różnych genów. Jest to więcej niż u typowego wirusa ale o wiele mniej niż 2556 domniemanych sekwencji kodujących białko znalezionych w pandorawirusach. Tak więc jego genom jest znacznie mniej gęsto upakowany niż w jakimkolwiek innym znanym wirusie. Dwie trzecie jego białek różnią się ponadto od białek innych wirusów. Pomimo fizycznego podobieństwa do pandorawirusa, sekwencja genomu Pithovirusa wykazuje słaby związek z owym wirusem, a bardziej przypomina przedstawicieli rodzin Marseillevindae, Megaviridae i Iridoviridae.

(...)
Wirusy nie są zdolne do samodzielnego rozmnażania się. W celu powielania własnych genów prowadzą proces namnażania, wykorzystując aparat kopiujący zawarty w żywych komórkach. Mogą zakażać wszystkie typy organizmów od zwierząt i roślin po bakterie i archeony. Dany gatunek wirusa zawiera tylko jeden rodzaj kwasu nukleinowego (DNA lub RNA), chociaż w trakcie rozwoju wewnątrz komórki dochodzi zwykle do syntezy drugiego rodzaju kwasu.
Ze względu na pasożytnictwo komórkowe wirusy posiadają na swojej powierzchni białka, które pozwalają zaatakować odpowiednie komórki. Wirusy nie posiadają rybosomów. Poza komórką nie wykazują żadnego metabolizmu, nie są zdolne do wzrostu ani rozmnażania się. Można je krystalizować.
Budowa wirusów: Zagadnienia dotyczące budowy wirusów dotuczą właściwie tylko stadium zdolnego do zakażenia komórek gospodarza. Pojedynczą, aktywną jednostkę wirusa nazywamy wirionem. Każdy wirion wykazuje obecność określonych elementów, a są nimi kapsyd i kwas nukleinowy.
- kapsyd = płaszcz białkowy - okrywa on kwas nukleinowy, zbudowany jest z łańcuchów białkowych, zwanych kapsomerami,
- kwas nukleinowy - niesie informację genetyczną niezbędną do replikacji oraz koduje białka strukturalne (kapsomery), ewentualnie enzymy (np. odwrotną transkryptazę).
Kwas nukleinowy razem z kapsydem nazywamy nukleokapsydem, czyli białkiem złożonym w którym grupę prostetyczną stanowi kwas nukleinowy.
Oprócz tego niektóre wirusy mogą być dodatkowo otoczone osłonką lipidową. Dotyczy to tych serotypów (Serotyp = odmiana mikroorganizmu, którą można określić za pomocą reakcji serologicznych, czyli reakcji z użyciem przeciwciał lub dopełniacza - różnice między serotypami zależą od antygenów znajdujących się na powierzchni komórek drobnoustroju), które uwalniają się z komórki przez pączkowanie. Ponieważ błona jest im zwykle potrzebna do kolejnej infekcji, takie wiriony są wrażliwe na układ dopełniacza je niszczący.


Istotną cechą systematyczną jest zagadnienie symetrii wirionu. Wyróżnia się jej trzy rodzaje:
- symetrię kubiczną - wirion ma kształt bryły foremnej, zwykle jest to dwudziestościan foremny (ikozaedr), stąd pochodzi inna nazwa tej symetrii - symetria ikozaedralna,
- symetrię helikalną - obserwujemy ją u wirusów mających śrubowato zawinięty nukleokapsyd,
- symetrię złożoną - opisuje wirusy niedające się zaliczyć do dwóch poprzednich rodzajów symetrii.
Symetria wirionu może nie być dostrzegalna na pierwszy rzut oka, co wynika z faktu istnienia osłonek lipidowych, mogących zakrywać rzeczywisty kształt nukleokapsydu. Jest tak zwłaszcza w przypadku wirusów o symetrii helikalnej, których otoczka jest dodatkowo wzmocniona warstwą tzw. białka M.


Strategie replikacyjne wirusów: Namnażanie wirusów jest zależne od rodzaju kwasu nukleinowego, który znajduje się w wirionie.
- Wirusy zawierające dsDNA: W tym przypadku wirus po wniknięciu do komórki rozpoczyna najpierw wytworzenie tzw. "wczesnego mRNA" na matrycy DNA pochodzącego z wirionu. Zwykle jednym z genów zawartych w genomie wirusa i odczytywanym poprzez wczesne mRNA jest DNA-zależna polimeraza DNA, która dokonuje powielenia wirusowego DNA. Dopiero z takich powielonych cząsteczek DNA następuje produkcja "późnego mRNA", kodującego kapsomery oraz inne białka uczestniczące w składaniu wirionów.
- Wirusy zawierające ssDNA: Mogą one zawierać w kapsydach zarówno DNA o polarności dodatniej, jak i ujemnej. Charakterystyczną cechą jest pętelka powstała przez zawinięcie końcówki liniowej cząsteczki DNA, która służy jako starter podczas replikacji DNA. Sam proces replikacyjny jest stosunkowo skomplikowany i wiąże się z kilkukrotną replikacją materiału genetycznego, który następnie jest rozdzielany na nici i cięty za pomocą nukleaz.
- Wirusy ssRNA o dodatniej polarności: Po wniknięciu wirusa do komórki następuje produkcja białek bezpośrednio z genomowego RNA, który może w tym wypadku pełnić rolę mRNA. W pewnym momencie rozwoju, gdy wytworzone zostaną odpowiednie białka, następuje powstanie dwuniciowej formy pośredniej RNA, która składa się zarówno z macierzystej nici o dodatniej polarności, jak i z nici o polarności ujemnej. Ta właśnie nić jest matrycą dla produkcji wielu kopii genomowego ssRNA(+).
- Wirusy ssRNA o ujemnej polarności: w ich przypadku musi najpierw dojść do stworzenia kopii o charakterze mRNA (czyli RNA o dodatniej polarności), a następnie z tych kopii są produkowane białka. Powstaje też dwuniciowa forma pośrednia, której nić RNA(+) służy do powstania licznych genomów RNA (-).
- Retrowirusy: Synteza kwasów nukleinowych u retrowirusów jest dosyć skomplikowana, ale jej kluczowym etapem jest zawsze przepisanie wirusowego RNA na DNA za pomocą obecnej w wirionie odwrotnej transkryptazy. Powstały w ten sposób ssDNA jest uzupełniany o drugą nić, zaś nowo utworzony dsDNA wbudowuje się w genom gospodarza, stanowiąc tzw. prowirus. DNA prowirusa służy zarówno do wytworzenia mRNA dla białek wirusowych, jak i dla wytworzenia potomnych genomów RNA.
OPIS INFEKCJI WIRUSOWEJ: Zakażenie komórki (z łac. infectio) przez wirusy może przebiegać - w zależności od gatunku - na wiele sposobów. Jednakże niezależnie od występujących różnic, podstawowe procesy zachodzące podczas takiej infekcji są wspólne dla wszystkich wirusów. Najbardziej ogólny schemat przedstawiony jest na poniższym rysunku:

Znaczenie poszczególnych etapów przedstawia się następująco:
- Adsorpcja - jest to proces przylegania wirusa do powierzchni komórki - jest oczywiście niezbędnym wstępem do zakażenia. Opiera się ona na połączeniu ze swoistym receptorem, z czego z kolei wynika tropizm tkankowy wirusa. Białko wirusowe, od którego zależy rozpoznanie komórki, to białko wiążące receptor. [Tropizm tkankowy to zjawisko polegające na selektywnym wyborze tkanki przez wirusa, które jest przyczyną atakowania określonych komórek przez ten rodzaj patogenów. Tropizm tkankowy wynika z faktu istnienia białek wiążących specyficzne dla danej tkanki receptory komórkowe. Przykładowo, poliowirus atakuje tkankę nerwową, w szczególności komórki rogów przednich rdzenia kręgowego odpowiedzialne za unerwienie mięśni, ale nie wykazuje powinowactwa do tkanki mięśniowe].
- Penetracja - jest ona procesem wnikania wirusa do komórki po jego uprzednim połączeniu się z receptorem. Może ono zachodzić na jeden z trzech podstawowych sposobów:
- fuzja - zachodzi w przypadku wirusów, które są otoczone błoną lipidową zawierającą białko fuzyjne. Otoczka lipidowa wirusa zlewa się z błoną komórkową, dzięki czemu wirus wnika do wnętrza.
- wiropeksja - jest ona sposobem polegającym na wykorzystaniu naturalnych mechanizmów komórki, które są wykorzystywane do pobierania różnych substancji odżywczych i regulacyjnych. Także w tym przypadku wirus musi posiadać otoczkę, gdyż na jednym z etapów wiropeksji dochodzi do zlewania się błon.

- "wślizgiwanie się" (endocytoza) - polega na bezpośrednim przejściu przez błonę komórki. Zachodzi ono w przypadku wirusów bezotoczkowych.
- Odpłaszczenie wirusa polega na uwolnieniu materiału genetycznego wirusa. W przypadku fuzji i wiropeksji zwykle następuje ono już podczas wnikania, gdyż jest bezpośrednio związane z mechanizmem penetracji.
- Produkcja białek wczesnych - zanim genom zostanie zreplikowany, często zdarza się, że potrzebne są białka niezbędne do pewnych czynności z tym związanych oraz inne, odpowiedzialne za zmianę metabolizmu komórki.
- Replikacja genomu - zachodzi w różny sposób, zależnie od charakteru materiału genetycznego, co zostało przedstawione wcześniej. Tutaj może dojść także do integracji genomu wirusa z genomem gospodarza.
- Produkcja białek późnych - zachodzi z reguły na podstawie kodu genetycznego ze świeżo wyprodukowanych nowych genomów. Są to zwykle białka strukturalne, wchodzące w skład kapsydu, oraz białka umożliwiającego prawidłowe złożenie wirionów.
- Składanie wirionów - jest to proces, w którym dochodzi do wytworzenia nukleokapsydów.
- Uwalnianie wirionów z komórki - następuje po ich złożeniu. Wirusy bezotoczkowe zwykle uwalniają się po śmierci komórki i jej rozpadzie, natomiast wirusy otoczkowe pączkują z powierzchni komórki. Otoczka lipidowa wirusa to zwykle pozyskany na tym etapie fragment błony komórkowej gospodarza.

Znaczenie medyczne: Według medycyny akademickiej wirusy wywołują niektóre choroby. Leczenie infekcji wirusowej jest trudne, gdyż wirusy nie posiadają własnego metabolizmu, który można by zablokować, jak to czyni się za pomocą antybiotyków w przypadku bakterii. Większość metod polega na ograniczeniu dalszego rozwoju infekcji, co przekształca chorobę wirusową w chorobę przewlekłą. Sytuację komplikuje fakt, iż zakażeniom wirusowym mogą towarzyszyć zakażenia bakteryjne. Zakażenia mogą nawracać (latencja wirusów) lub też ujawniać się po bardzo długim czasie. Wiele chorób wirusowych jest nadal nieuleczalnych (np. wścieklizna, AIDS). Ponadto wirusy onkogenne związane są z rozwojem chorób nowotworowych.



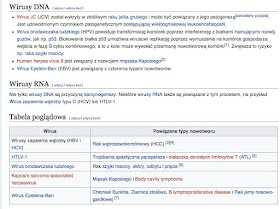

(...)
Leki przeciwwirusowe można podzielić na:
- substancje wzmacniające układ odpornościowy w walce z wirusami - na przykład interferon w leczeniu wirusowego zapalenia wątroby,
- substancje blokujące rozwój wirusa, na przykład poprzez blokadę białek wirusowych,
- substancje blokujące białka receptorowe, uniemożliwiające przyłączanie wirusów do błony komórkowej
Jedną z metod walki z chorobami wirusowymi są szczepienia.

Dość duże grupy wirusów dają się łączyć w monofiletyczne taksony, jednak nie ma póki co dowodów, żeby wszystkie one pochodziły od jednego prawirusa.
Klasyfikacja wirusów: Opracowaniem systematyki wirusów zajmuje się Międzynarodowy Komitet Taksonomii Wirusów, jednak ze względu na niewielką ilość danych dotyczących filogenezy i pokrewieństwa jest ona dosyć płynna. Różne ośrodki naukowe zwykle różnią się, zwłaszcza w szczegółach, co do klasyfikacji wirusów.
Wirusy dzieli się na zwierzęce i roślinne albo, ze względu na ich wielkość, na:
- wirusy duże (150–300 nm)
- pokswirusy
- wirusy średnie (50–150 nm)
- herpeswirusy
- adenowirusy
- wirusy małe (zwierzęce 20–50 nm)
- pikornawirusy
- parwowirusy

Ze względu na organizację materiału genetycznego dzieli się wirusy na:
- wirusy RNA (w tym retrowirusy)
- wirusy DNA.
Niektóre dotychczas poznane wirusy chorobotwórcze to:
- wirus grypy
- wirus zespołu nabytego braku odporności (HIV)
- wirus choroby Heinego-Medina
- wirus choroby skokowej owiec
- wirus Ebola
- wirus dengi
- wirus opryszczki pospolitej
- wirus odry
- wirus zapalenia wątroby typu A, wirus zapalenia wątroby typu B, wirus zapalenia wątroby typu C
- wirus brodawczaka ludzkiego.
Wirofagi: Wirofagi to wirusy potrzebujące do replikacji obecności innych wirusów (wirusy satelitarne) i hamujące ich replikacje. Pierwszy odkryty wirus o takich właściwościach nazwano sputnik. Kolejne to: CroV oraz Organic Lake Virophage (OLV).
 Wirofagami zwiemy wirusy niezdolne do samodzielnej replikacji. Stanowią one podgrupę wirusów satelitarnych. Rozwój wirofaga wymaga koinfekcji komórki przez wirofaga i innego wirusa. Po rozpoczęciu namnażania wirusa-ofiary wirofag atakuje go, wykorzystując białka i materiał genetyczny do replikacji własnego wirionu. Do tej pory poznanymi wirofagami są: Sputnik, Mavirus, OLV (Organic Lake Virophage), Zamilon. Są to wirusy DNA. Wirofag Zamilon atakuje wirusy Mimivirus, pasożytujące na komórkach ameb Acanthoamoeba polyphaga. W 2016 roku odkryto, że jeden ze szczepów mimiwirusów wykazuje odporność na zakażenie tym wirofagiem. System MIMIVIRE (ang. Mimivirus Virophage Resistance Element) wykorzystuje mechanizm podobny do systemu CRISPR (ang. Clustered Regularly Interspaced Short Palindromic Repeats) występującego u bakterii i archeonów, który sprawdza DNA pod kątem obecności fragmentów z „biblioteki” obcych genów i tnie potencjalnie niebezpieczną nić. Potwierdzeniem tej hipotezy jest fakt, że wyciszenie tego regionu kodu genetycznego mimiwirusa sprawia, że staje się on podatny na atak wirofaga.
Wirofagami zwiemy wirusy niezdolne do samodzielnej replikacji. Stanowią one podgrupę wirusów satelitarnych. Rozwój wirofaga wymaga koinfekcji komórki przez wirofaga i innego wirusa. Po rozpoczęciu namnażania wirusa-ofiary wirofag atakuje go, wykorzystując białka i materiał genetyczny do replikacji własnego wirionu. Do tej pory poznanymi wirofagami są: Sputnik, Mavirus, OLV (Organic Lake Virophage), Zamilon. Są to wirusy DNA. Wirofag Zamilon atakuje wirusy Mimivirus, pasożytujące na komórkach ameb Acanthoamoeba polyphaga. W 2016 roku odkryto, że jeden ze szczepów mimiwirusów wykazuje odporność na zakażenie tym wirofagiem. System MIMIVIRE (ang. Mimivirus Virophage Resistance Element) wykorzystuje mechanizm podobny do systemu CRISPR (ang. Clustered Regularly Interspaced Short Palindromic Repeats) występującego u bakterii i archeonów, który sprawdza DNA pod kątem obecności fragmentów z „biblioteki” obcych genów i tnie potencjalnie niebezpieczną nić. Potwierdzeniem tej hipotezy jest fakt, że wyciszenie tego regionu kodu genetycznego mimiwirusa sprawia, że staje się on podatny na atak wirofaga.
Wirusy satelitarne - są to wirusy, których replikacja jest zależna od innych wirusów. Mogę one kodo-
-wać własne białka albo opierać się na innych wirusach zarówno dla enkapsydacji, jak i replikacji. Pie-
-rwszy wirus satelitarny opisano w 1962r., był on powiązany z wirusem nekrozy tytoniu (TNV, ang.
Tobacco necrosis virus). Niektóre wirusy satelitarne szkodzą wirusom-gospodarzom lub uniemożliwia-
-ją ich rozwój; są one nazywane wirofagami.
W celu poszerzenia wiedzy na temat wirofagów oraz wirusów satelitarnych zachęcam do zapozna-
-nia się z pracą "WIROFAGI - NOWE ELEMENTY BIOLOGICZNE" dostępną na ResearchGate,
przygotowaną przez zespół naukowców z Katedry Immunologii, Wydziału Biologii Uniwersytetu
Szczecińskiego:

Bibliografia i wykorzystane źródła:
- Michał Skawiński: „System immunologiczny” wirusów obroną przed... innymi wirusami. DolinaBiotechnologiczna.pl, 2016-03-19. [dostęp 2016-04-23].
- Anthony Levasseur i inni, MIMIVIRE is a defence system in mimivirus that confers resistance to virophage, „Nature”, 531, 2016, s. 249–252, DOI: 10.1038/nature17146 (ang.).
- Olga Orzyłowska-Śliwińska, Wiadomo, że wirusy atakują bakterie, ale czy mogą atakować inne wirusy?. „Świat Nauki”. nr. 6 (238), s. 83, czerwiec 2011. Prószyński Media. ISSN 0867-6380
- Ed Yong: Giant virus resurrected from 30,000-year-old ice(ang.). W: Nature: News [on-line]. 2014-03-04. [dostęp 2014-03-04].
- Matthieu Legendre, Julia Bartoli, Lyubov Shmakova, Sandra Jeudy, Karine Labadie, Annie Adrait, Magali Lescot, Olivier Poirot, Lionel Bertaux, Christophe Bruley, Yohann Couté, Elizaveta Rivkina, Chantal Abergel, Jean-Michel Claverie. Thirty-thousand-year-old distant relative of giant icosahedral DNA viruses with a pandoravirus morphology. „Proceedings of the National Academy of Sciences”, 2014. Stanford University's HighWire Press. DOI: 10.1073/pnas.1320670111. ISSN 1091-6490 (ang.).
- Marilyn J. Roossinck, David Sleat, Peter Palukaitis. Satellite RNAs of plant viruses: structures and biological effects. „Microbiological reviews”. 56 (2), s. 265-279, 1992.
- R.I.B. Francki. Plant virus satellites. „Annu. Rev. Microbiol.”. 39, s. 151-174, 1985. DOI: 10.1146/annurev.mi.39.100185.001055.
- J.M. Kaper: Modulation of viral plant diseases by secondary RNA agents. W: RNA genetics. E. Domingo, J.J. Holland, P. Ahlquist (red.). T. III. Variability in RNA genomes. Boca Raton, Floryda, USA: CRC Press, Inc., 1988, s. 171-194. ISBN 0-8493-6668-2.
- A.F. Murant. Satellites of plant viruses. „Annu. Rev. Phytopathol.”. 20, s. 49-70, 1982. DOI: 10.1146/annurev.py.20.090182.000405.
- A.E. Simon. Satellite RNAs of plant viruses. „Plant Mol. Biol. Rep.”. 6, s. 240-252, 1988. DOI: 10.1007/BF02670384 B. Kassanis. Properties and behaviour of a virus depending for its multiplication on another. „J. Gen. Microbiol.”. 27, s. 477-488, 1962. DOI: 10.1099/00221287-27-3-477
Brak komentarzy:
Prześlij komentarz